Lūksne
Šajā rakstā ir pārāk maz vikisaišu. Lūdzu, palīdzi uzlabot šo rakstu, saliekot tajā saites uz citiem rakstiem. Ja ir kādi ieteikumi, vari tos pievienot diskusijā. Vairāk lasi lietošanas pamācībā. |
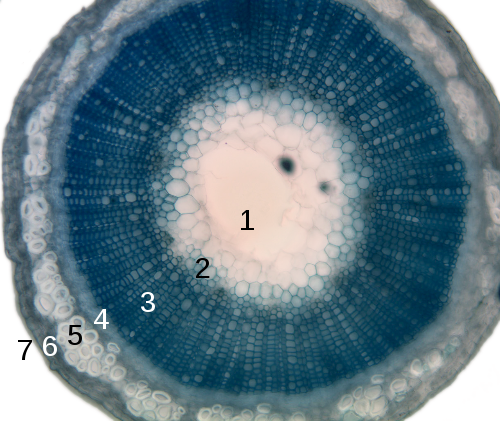
1. Serde,
2. Protoksilēma,
3. Koksne I,
4. Lūksne I,
5. Sklerenhīma,
6. Miza,
7. Epiderma
Lūksne jeb floēma ir vaskulāro augu audu komplekss, kas pārvada organiskās vielas (fotosintēzes produktus), jo īpaši saharozi,[1] cukuru, uz auga daļām. Kokos lūksne ir visdziļākais mizas slānis, tādēļ vārds 'floēma', atvasināts no grieķu vārda φλοιός (phloios) nozīmē "korķis".
Uzbūve
[labot šo sadaļu | labot pirmkodu]Lūksnes audi galvenokārt sastāv no divu veidu šūnām bez sekundārajiem šūnapvalkiem: sietstobriem un pavadītājšūnām (vai albumīnšūnām kailsēkļiem). Lūksne iekļauj arī parenhīmas šūnas un atbalsta šūnas, tādas kā šķiedras un sklereīdas.
Pavadītājšūnas ar sietstobriem
[labot šo sadaļu | labot pirmkodu]
1. Koksne
2. Lūksne
3. Kambijs
4. Serde
5. Pavadītājšūnas
Sietstobri ir šūnu veids, kas ir atbildīgi par cukuru transportu viscauri augam.[2] Briedumā tiem trūkst kodola un ir tikai dažas organellas, tādēļ tie paļaujas uz pavadītājšūnām jeb albumīnšūnām priekš visām savām metaboliskajām vajadzībām. Sietstobru šūnas satur vakuolas un citas organellas, piemēram, ribosomas pirms tās nobriest, bet tās pamatā migrē uz šūnapvalku un izšķīst briedumā; tas nodrošina mazāku kavēšanos šķidrumu kustībā. Viena no dažām organellām, ko satur sietstobri briedumā, ir graudainais endoplazmatiskais tīkls, kas var atrasties plazmatiskajā membrānā, bieži vien, blakus plazmodesmām. Poras tiek pastiprinātas ar polisaharīdu trombiem sauktiem par callose.[2]
Parenhīmas šūnas
[labot šo sadaļu | labot pirmkodu]Pavadītājšūnas
[labot šo sadaļu | labot pirmkodu]Sietstobru metaboliskās funkcijas ir atkarīgas no ciešas sadarbības ar pavadītājšūnām, specializētas formas parenhīmas šūnām. Visas sietstobru šūnas pamatfunkcijas ir pārnestas uz (daudz mazāko) pavadītājšūnu, tipisku kodolsaturošu augu šūnu, izņemot to, ka pavadītājšūnām parasti ir lielāks skaits ar ribosomām un mitohondrijiem. Pavadītājšūnas blīvā citoplazma ir savienota ar sietstobra šūnu ar plazmodesmām.[2] Kopējā starppārrāvumā, ko dala sietstobra elements un pavadītājšūna, ir daudz plasmodesmas, kas veido stobra laukumu. Ir divu veidu pavadītājšūnas.
- Parastās pavadītājšūnas, kurām ir plānas sieniņas un ir dažas vai nav plasmodesmu savienojuma ar citām šūnām, izņemot sietstobru.
- Pārvietošanas pavadītājšūnas, kurām ir ļoti krokotas sieniņas, kas ir blakus nesietstobru šūnā, dodot vairāk vietas pārvietošanai. Tās ir specializējušās atkritumproduktu savākšanā no šūnu sieniņām, kuras aktīvi pumpē pieprasot enerģiju.
Albumīnšūnas
[labot šo sadaļu | labot pirmkodu]Albumīnšūnām ir līdzīga loma kā pavadītājšūnām, bet tās ir saistītas tikai ar sietstobriem un ir atrodamas tikai bezsēklu vaskulārajos augos un kailsēkļos.[2]
Citas parenhīmas šūnas
[labot šo sadaļu | labot pirmkodu]Citas parenhīmas šūnas lūksnē pamatā nav diferencētas un tiek izmantotas barības vielu uzglabāšanai.[2]
Balstaudu šūnas
[labot šo sadaļu | labot pirmkodu]Lai gan lūksnes primārā funkcija ir transportēt cukurs, tā arī satur šūnas, kas veic mehāniskā atbalsta funkciju. Tās galvenokārt iedala divās kategorijās: šķiedrās un sklereīdās. Abiem šūnu veidiem ir dubulti šūnapvalki, kas ir izraisījis miršanu briedumā. Dubultais šūnapvalks palielina to stingrību un stiepes deformācijas stiprumu.
Šķiedras
[labot šo sadaļu | labot pirmkodu]Šķiedras ir garas, šauras atbalsta šūnas kas nodrošina vilkšanas spēku bez elastības ierobežošanas. Tās ir arī atrodamas koksnē un ir galvenā sastāvdaļa dažādos tekstilmateriālos, piemēram, papīrā, linā un kokvilnā.[2]
Sklereīdas jeb akmensšūnas
[labot šo sadaļu | labot pirmkodu]Sklereīdas (akmensšūnas) ir nedzīvas, neregulāras formas šūnas ar pārkoksnējušos šūnapvalku, kas palielina saspiešanas spēku[2], bet var samazināt elastību dažādās pakāpēs. Tās kalpo aizsardzībā pret zālēdājiem, jo to neregulārās forma un stiprums palielina slodzi uz zobiem, kad zālēdāji košļā. Piemēram, tās ir atbildīgas par grantaino struktūru bumbieros.
Funkcijas
[labot šo sadaļu | labot pirmkodu]Atšķirībā no koksnes (kas pamatā sastāv no atmirušām šūnām), lūksne sastāv no mierīgi dzīvojošām šūnām, kas transportē sulu. Sula ir ūdens šķīdums, kas bagātināts ar cukuriem no fotosintēzes galaproduktiem. Šīs vielas tiek transportētas uz auga nefotosintezējošām daļām, piemēram, saknēm, vai uz uzglabāšanas vietām kā gumi vai sīpoli.
Ernst Münch 1930. gadā ierosināja spiediena plūsmas hipotēzi, kas izskaidroja lūksnes vadīšanas mehānismu.[3]
Auga augšanas periodā, parasti pavasarī, barības vielu uzkrājēj-orgāni, piemēram, saknes ir cukura avoti, un daudzas augu augšanas vietas ir kā cukura krātuves. Kustība lūksnē notiek uz visām pusēm, bet koksnes šūnās tā ir vienvirziena (augšup).
Pēc augšanas perioda, kad meristēma ir pārstājusi darboties, lapas ir barības vielu avoti un uzkrājēj-orgāni ir krātuves. Attīstībā esošie sēklu nesējorgāni (piemēram, augļi) vienmēr ir krātuves. Dēļ daudzvirzienu plūsmas, kā arī dēļ fakta, ka sula nevar kustēties starp blakusesošiem sietstobriem, nav nekas neparasts, ka sula blakusesošos sietstobros plūst pretējos virzienos.
Kamēr koksnē ūdens un minerālu kustība lielākoties pārvietojas ar negatīvu spiedienu (spriegumu), kustība caur lūksnes vadaudiem notiek ar pozitīvu hidrostatisku spiedienu. Šo procesu sauc par pārvietošanu, un tas tiek paveikts ar procesiem, ko sauc lūksnes piepildīšanu un iztukšošanu. Šūnas cukura avotos ar aktīvā transporta palīdzību "pielādē" sietstobru šūnas ar izšķīdušajām molekulām. Tas izraisa osmozi — ūdens ieplūšanu šūnās, radot spiedienu, kas spiež sulu lejup pa vadaudiem. Cukura krātuvēs, šūnas aktīvi transportē šķīdumus ārā no sietstobriem, radot pilnīgi pretēju efektu.
Tomēr izrādās, ka daži augi ar aktīvo transportu nepiepilda lūksni. Šādi gadījumi zināmi kā polimēru slazda mehānisms, ko aprakstījis Roberts Turgeons.[4] Šādos gadījumos mazie cukuri, kā piemēram saharoze, ieplūst starpšūnās caur plānajām plasmodesmām, kur tie polimerizējas par rafinozi (trisaharīds) un citiem lielākiem oligosaharīdiem. Tagad tie nav spējīgi pārvietoties atpakaļ, bet var doties cauri plānākām plasmodesmām uz sietstobriem.
Spiediena plūsmas hipotēze skaidro lūksnes sulas transporta mehānismu,[5] lai gan ir bijušas arī citas skaidrojošas hipotēzes.[6] Lūksnes sula spēlē arī svarīgu loma kā informatīvo signālu sūtītāja viscaur vaskulārajiem augiem. "Piepildīšanas un iztukšošanas modeli lielākoties nosaka vadītspēja un plasmodesmu skaits, un plazmas membrānas transporta proteīni, kuru funkcijas ir atkarīgas no pozīcijas.Nesenie pētījumi liecina ka mobilie proteīni un RNS ir daļa no auga lielas-distances komunikācijas signalizēšanas sistēmas. Eksistē arī pierādījumi par tiešu makromolekulu transportēšanu un kārtošanu, kad tās iziet cauri plasmodesmām."[6]
Simplastiskā lūksnes piepildīšana (iepriekšminētais polimēru slazda mehānisms) ir lielākoties tropu lietus mežu augiem un tiek uzskatīts par primitīvāku mehānismu. Aktīvais transports, kas aploplastiski piepilda lūksni, tiek apskatīts kā daudz sarežģītāks mehānisms, kas ir atrodams vēlāk-evolucionējušos augos, it īpaši, mērenajos un sausajos klimata apstākļos. Šis mehānisms tādēļ atļauj augiem kolonizēties aukstākās vietās.
Organiskās molekulas kā cukuri, aminoskābes, noteikti hormoni un pat RNS ziņotāji tiek transportēti lūksnē caur sietstobru šūnām.[6]
Joslojums
[labot šo sadaļu | labot pirmkodu]Lūksnes vadaudi atrodas ārpus koksnes lielākajā daļā augu, tāpēc kokus vai citus augus var efektīvi iznīcināt noraujot nost gredzena mizu stumbram vai stublājam. Ja lūksne ir iznīcināta, barības vielas nevar sasniegt saknes un koks/augs nomirs. Koki, kas atrodas tādās vietās, kur dzīvo dzīvnieki kā bebri, ir viegli ievainojami, jo bebri nograuž mizu ļoti precīzā augstumā. Šo procesu sauc par joslošanu un to var izmantot agrokulturāliem nolūkiem. Piemēram, milzīgie augļi un dārzeņi, ko var redzēt tirdziņos un karnevālos, ir audzēti joslojot. Zemkopis izgriež joslu no mizas pie liela zara pamatnes un nogriež visus, izņemot vienu augli/dārzeni no tā zara. Tādā veidā, visiem cukuriem, ko saražos lapas no tā zara nebūs kur uzkrāties (uzkrājēj-orgāni) kā tikai šajā vienā auglī/dārzenī, kas pārsniegs vairākas reizes savus normālos izmērus.
Veidošanās
[labot šo sadaļu | labot pirmkodu]Kad augs ir dīgļa stadijā, vadaudi rodas no prokambija audiem, kas ir dīgļa centrā, prokambija šūnām daloties un diferencējoties. Protofloēma pati par sevi parādās vidusdzīslā iespiežoties dīgļlapu izaugumā, kas ir pirmā lapas attīstības stadija segsēkļiem. Hormons auksīns, ko transportē proteīns PIN1, ir atbildīgs par protofloēmas šķiedru augšanu, signalizē par audu pēdējo stāvokli. SHORTROOT(SHR) un microRNS165/166 arī piedalās šajā procesā, kamēr Kallozes sintēze 3 (CALS3) nomāc vietu, kur SHORTROOT (SHR) un microRNS165 vajag doties.
Dīglī saknes lūksne attīstās patstāvīgi augšējā hipokotilā, kas atrodas blakus dīgļsaknei un dīgļlapai.[7]
Pieaugušiem augiem lūksnes audi veidojas izaugot no meristēmas šūnām kambijā. Lūksnes veidošanos izdala fāzēs. Primārajā fāzē lūksnes attīstību nosaka apikālā meristēma un tā veidojas no prokambija. Sekundārajā fāzē vaskulārais kambijs nosaka lūksnes veidošanos uz āru uz iekšienē jau esošajiem lūksnes slāņiem.
Dažās divdīgļlapju dzimtās (Apocynaceae, Convolvulaceae, Cucurbitaceae, Solanaceae, Myrtaceae, Asteraceae) lūksne veidojas arī uz iekšpusi no kambija; šajā gadījumā šī atšķirība starp ārējo lūksni un iekšējo lūksni rodas. Iekšējā lūksne ir lielākoties primārā un sāk diferencēšanos vēlāk kā ārējā floēma un protofloēma, lai gan tas ir ar izņēmumiem. Citās dzimtās (Amaranthaceae, Nyctaginaceae, Salvadoraceae) arī kambijs periodiski veidojas iekšpusē lūkšņu šķiedrām: šādas lūkšņu šķiedras sauc par iekļauto lūksni.[8]
Lietošana uzturā
[labot šo sadaļu | labot pirmkodu]
Priedes lūksne tiek lietota Somijā kā ēdiena aizstājējs bada laikos un pat labajos gados ziemeļaustrumos. Lūksnes krājumi no iepriekšējiem gadiem palīdzēja nenomirt badā lielajā badā 1860. gados. Lūksne tiek izžāvēta un sasmalcināta līdz miltiem (pettu somiski) un sajaukti ar rudziem izveidojot cietu tumšu maizi, mizas maizi. Vismazāk novērtētais bija silkko, maize veidota tikai no paniņām un pettu bez jebkādiem īstiem rudziem vai labības graudu miltiem. Nesen pettu ir nācis atkal modē dēļ ziņkārības un daži apgalvo, ka tas ir ļoti veselīgs. Tomēr, uzturvērtības ziņā tā enerģētiskā vērtība ir ļoti zema salīdzinot ar rudziem vai citiem graudiem.
Lūksne no Āra bērza (Nokarenais bērzs, Kārpainais bērzs) arī ir tikusi izmantota miltu veidošanai pagātnē.
Atsauces
[labot šo sadaļu | labot pirmkodu]- ↑ Lalonde S. Wipf D., Frommer W.B. (2004). "Transport mechanisms for organic forms of carbon and nitrogen between source and sink". Annu Rev Plant Biol. 55: 341–72. doi:10.1146/annurev.arplant.55.031903.141758. PMID 15377224.
- ↑ 2,0 2,1 2,2 2,3 2,4 2,5 2,6 Peter H. Raven, Ever, R.F., Eichhorn, S.E. Biology of Plants. New York, NY, U.S.A. : Worth Publishers, 1992. 791. lpp.
- ↑ E Münch. Die Stoffbewegunen in der Pflanze. Verlag von Gustav Fischer, Jena, 1930. 234. lpp.
- ↑ Turgeon, R. «Symplastic phloem loading and the sink-source transition in leaves: a model». In VL Bonnemain, S Delrot, J Dainty, WJ Lucas, (eds). Recent Advances Phloem Transport and Assimilate Compartmentation, 1991. ISBN 2908261618.
- ↑ Khan, Aslam. Plant Anatomy And Physiology. Gyan Publishing House, 2001. gada 1. janvāris. ISBN 978-81-7835-049-3. Skatīts: 2013. gada 6. aprīlis.
- ↑ 6,0 6,1 6,2 Turgeon, Robert; Wolf, Shmuel (2009). "Phloem Transport: Cellular Pathways and Molecular Trafficking". Annual Review of Plant Biology 60: 207–21. doi:10.1146/annurev.arplant.043008.092045. PMID 19025382.
- ↑ Lucas, William, et al. The Plant Vascular System: Evolution, Development and Functions. Journal of Integrative Plant Biology. 55, 294-388 (2013) PMID 23462277
- ↑ Evert, Ray F. Esau's Plant Anatomy. John Wiley & Sons, Inc, 2006, pp. 357—358, ISBN 0470047372.
|