神經編碼


神經編碼(粵拼:san4 ging1 pin1 maa5;英文:neural coding)係神經科學嘅一個子領域,專門研究神經細胞(neuron)點樣對外界刺激有反應,以及神經細胞射嗰啲訊號點樣表達佢哋所帶嗰啲資訊嘅內容:一隻脊椎動物嘅感官會無時無刻噉對外界刺激起反應-眼會感應光、耳仔會感應聲,呢啲感官嘅感覺神經細胞會向個腦射訊號,個腦跟住會對啲訊號作出處理,並且射返啲(同個腦收到嗰啲訊號唔同樣嘅)新訊號落去身體嘅各部份嗰度,教身體嘅肌肉同腺作出適當嘅反應[1][2]。
喺呢個過程當中,神經系統做咗將訊號轉化嘅工作,即係話一粒神經細胞射嘅訊號實會同佢所受嘅刺激成某啲關係:定義上,一個訊號實係會同佢所測量嗰個物理量成特定嘅函數(簡單講即係成一個特定嘅數學關係),所以睇訊號嘅嗰方先有可能知道個訊號係傳達緊啲乜嘢資訊[3][4];舉個例說明,一粒感光細胞(視網膜上對光起反應嘅細胞)射嘅訊號嘅物理特性 (例如係頻率同時間差)一定係同佢感應到嘅光嘅某啲物理特性 (例如光嘅亮度或者色水)成特定嘅關係,否則接收訊號嗰一方(個腦)唔會有可能憑訊號嘅物理特性解讀訊號傳嘅資訊[5]。
神經編碼想做嘅嘢大致上就係想拆解神經系統嘅呢啲函數:研究神經編碼嘅科學家會係噉做實驗,睇吓唔同嘅刺激會引起乜嘢規律嘅神經細胞訊號,靠噉嚟研究神經系統係點樣將資訊編碼(coding)嘅-即係想「將個腦解碼」[6][7];而呢啲研究所得嘅知識可以配合光遺傳學等嘅技術用嚟醫治神經性嘅疾病[8][9]。
理論基礎
[編輯]
神經細胞
[編輯]- 神經細胞(neuron)係一種與別不同嘅細胞,佈滿動物嘅神經系統:神經細胞分好多種,每種都會對某啲特定類型嘅刺激有反應;當一粒神經細胞受到佢相應嗰種刺激嗰陣,會沿由佢細胞體伸出嚟嗰條軸突(axon)射出叫做動作電位(action potential,或者 spike)嘅電脈衝;而好多神經細胞喺受到第啲神經細胞射嘅動作電位刺激嗰時,又會射新動作電位-所以當一粒神經細胞受刺激而射動作電位嗰陣時,好多時就會引致一連串嘅動作電位,令到訊號沿住神經系統傳開去,一路上到腦[10][11];舉個例說明,喺啲感覺器官嗰度嘅感覺神經細胞(sensory neuron)喺受到光、聲、氣味同溫度變化等嘅嘢刺激到嘅時候,都會射動作電位,而如果負責感應光嗰啲感覺神經細胞唔射動作電位,個腦就知道外界冇光[12]。
- 動作電位等神經細胞射出嚟嘅嘢係訊號(signal):廣義上,訊號可以理解為「是但一個可以傳達資訊嘅函數」(a function that conveys information)[13];想像兩個科研人員喺度研究太陽光,A 君攞住感應器去量度一束光嘅強度,佢哋駁好嗮電線,而 A 君同 B 君講好,佢會傳一個訊號,個訊號會係一個電壓,而電壓大細會同佢探測到嗰束光嘅強度成以下關係:;當中 係傳出去嗰個電壓, 係觀察到嗰束光嘅強度,而 係一個事先講好嘅數值-個訊號()同佢哋想傳嗰個資訊()成一個特定嘅函數,所以如果假設 A 君靠得住,B 君會能夠由呢個函數同收到嘅訊號當中,得知個資訊;動作電位都可以用同樣嘅道理嚟諗,一粒感光神經細胞射嘅動作電位嘅某啲物理特性(例如係射電位嘅頻率 )會同佢所受嘅刺激嘅特性(例如係束光嘅亮度 )成某一段特定嘅數學關係,,當中 係一個恆定嘅函數,所以個腦可以透過睇動作電位嘅特性知道外界刺激嘅特性[7]。
由簡單嘅觀察已知,腦有能力指揮身體對外界刺激俾適當嘅反應,所以腦接收到嘅訊號當中實係以某啲方法將資訊傳達咗俾個腦,而神經編碼想做嘅就係同呢啲訊號做解碼(decode)-研究神經編碼嘅科學家係想拆解神經訊號同所受嘅刺激成點樣嘅函數(啲 );理論上,如果具備足夠嘅神經編碼知識,一個人將會能夠齋靠睇神經訊號嘅規律,就知道嗰隻動物感知到啲乜同諗緊乜[14][15]。即係話神經編碼研究可以分做兩大門[2][16]:
- 神經入碼(neural encoding)研究外界刺激嘅物理特性(例:光嘅亮度同色水、聲嘅頻率、氣味反映嘅化學成份... 等等)同神經細胞射嘅訊號嘅物理特性之間有乜關係;同埋
- 神經解碼(neural decoding)研究點樣由睇到嘅神經訊號嗰度解讀返裏頭嗰啲資訊出嚟。
編碼方案
[編輯]
編碼方案(coding scheme)係神經編碼研究上嘅一個基本概念,一個編碼方案會表明一個訊號特性對應乜嘢資訊同點樣對應法,例:訊號嘅邊個特性對應乜嘢外界物理量,同埋個函數 係點嘅樣[17]。
一列(train)嘅動作電位可能會帶有多個唔同編碼理論排出嚟嘅資訊。舉個例說明,運動神經細胞係一種神經細胞,專門負責將訊號由腦傳去啲肌肉嗰度,指揮肌肉郁動;研究顯示,一嚿肌肉收縮得有幾勁係視乎運動神經細胞嘅發射頻率(firing rate;指每單位時間有幾多個動作電位)嘅,而同時每一個動作電位嘅「到達時間」又帶有一啲資訊喺入面,會控制肌肉活動嘅第啲部份-即係話有最少兩個編碼方案,第一個方案指明發射頻率同「嚿肌肉要收縮得有幾勁」有乜嘢關係,而第二個方案就指明到達時間同乜嘢資訊有關以及有點樣嘅關係[18]。因為噉,同一柞動作電位入面可以帶有好幾種唔同嘅資訊[19][20]。
研究方法
[編輯]神經編碼研究通常會用實驗嘅方法嚟做,會用一啲特定嘅方法量度動作電位(睇埋神經成像)之後,由量度攞到數據之後就郁手做統計分析,搵出外界刺激嘅特性會點樣影響到動作電位嘅種種特性[5][6]。
例如想像家陣有班研究者想研究視覺系統嘅神經編碼,想知「一個人感知到嘅光嘅色水會點樣影響動作電位嘅特性」,佢哋可以做好似以下噉嘅實驗:搵一班受試者返嚟,將佢哋分做幾組,每組都俾佢哋睇到一束光(刺激),但除咗光嘅色水之外,唔同組嘅受試者睇到嘅刺激完全一樣-所以任何組嘅受試者之間喺動作電位上嘅差異只可能係源於光嘅色水嘅差異[21][22];而譬如班研究者做完統計分析(可以睇吓比較平均值相關嘅嘢)之後發現,某柞感光神經細胞淨係會喺受到藍色光刺激到嗰陣以 5 Hz 嘅頻率射動作電位,而且淨係喺受到紅色光刺激到嗰陣先會以 10 Hz 嘅頻率射動作電位,噉佢哋喺下次見到嗰柞神經細胞以 5 Hz 嘅頻率射動作電位嗰陣,就可以唔使睇第啲嘢都知嗰個人見到藍色光[23]。
數學模型
[編輯]同任何科學上嘅研究一樣,神經編碼研究通常都要將動作電位想像成抽象化嘅數學物體。動作電位會引致神經細胞周圍嘅微電壓出現變化,而呢啲微電壓變化可以用好細支嘅電極量度,所以神經編碼研究要處理嘅原始數據(raw data;指直接量度到嘅數字)通常係一個表示電壓值隨時間變化嘅時間序列(time series);而當神經科學家要將動作電位想像成數學物體嗰陣,會考慮以下兩點:
- 動作電位嘅種種特性-例如係振幅、形狀、以及彼此之間嘅時間間隔[24]呀噉-係就係主要取決於個刺激,但會有些少隨機性嘅起伏,即係話就算兩個刺激完全一樣,兩者所分別引起嘅動作電位都會有些微嘅差異;
- 一個個別嘅動作電位短暫得好交關,通常唔會維持得耐過 1 毫秒(1 秒 = 1,000 毫秒);
所以喺實際研究上,研究者多數都會為咗分析上嘅方便,假設每一個動作電位都係一個板嘅,而且每一個動作電位所維持嘅時間極短(可以睇埋無窮小量同相關嘅數學概念)。於是喺多數嘅神經編碼嘅數學模型入面,一串動作電位就會俾人當係喺某一段時間之內一連串同樣大細、而且射完就喺剎那間消失嘅脈衝噉嚟睇[25]。
頻率編碼
[編輯]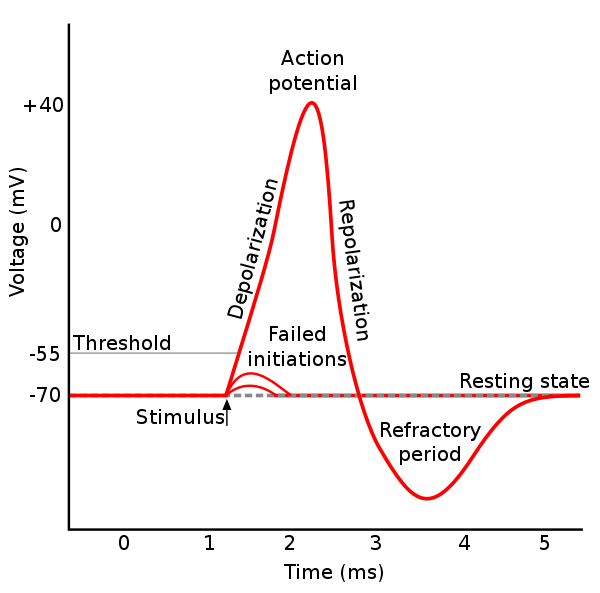
頻率編碼(rate coding / frequency coding)模型集中思考神經細胞射嗰啲 spike 嘅頻率(發射頻率;firing rate)當中帶啲乜資訊。頻率編碼响研究感官系統嘅神經編碼嗰陣好使得:喺感官系統入面,通常一個外界刺激嘅強度愈高(意思即係個刺激每單位面積嘅功率有幾高,強度高嘅光會掁眼,而強度高嘅聲會響),俾佢刺激到嗰啲神經細胞嘅發射頻率就會愈高[註 2][26]。
頻率編碼係神經科學上最先有人研究嘅編碼方案。最基礎嘅頻率編碼方案假設神經細胞傳嘅資訊大部份-甚至全部-都係記載咗喺啲 spike 嘅頻率入面嘅。呢類模型一般會假設神經細胞射 spike 呢個反應有些少隨機性,會用機會率數值嚟代表某一段指定時段(例如 1 毫秒)入面有 spike 出現嘅機會率,並且靠噉嚟計出所研究緊嗰柞神經細胞响某一段特定時間入面嘅發射頻率[註 3];喺呢個分析過程當中,啲模型會忽略啲 spike 嘅第啲時間性特徵(例如係到達時間)。研究顯示,呢種做法會忽略咗一啲重要資訊,但個好處係啲模型冇咁易受到「spike 之間嘅時間間隔」(inter-spike intervals,ISI)等嘅拉雜嘢干擾[24]。
神經發放率
[編輯]神經發放率(spike-count rate)係最簡單直接嗰種發射頻率計法。想像依家做一次試驗(trial;例如受試者受咗吓刺激),數吓嗰次試驗入面出現咗幾多個 spike,再將 spike 嘅數量除以嗰次試驗嘅時間長度[26]。
每一段攞嚟計發射頻率嘅時間間隔(time window;)都係由個研究者話事嘅,取決於量度緊嗰種神經細胞或者外界刺激嘅種類,典型嘅 通常都係喺 100 至 500 毫秒之間。舉個例說明:例如家吓做咗一次 5 秒(5,000 毫秒)長嘅試驗,喺嗰段試驗期間量度到 100 個喺時間上唔平均分佈嘅 spike,噉要計發射頻率有至少兩種做法-將發射頻率當做 (每毫秒 0.02 個 spike),又或者將次試驗嗰 5 秒時間斬件做 10 段各有 500 毫秒長嘅時間間隔(),計吓每一段間隔入面嘅 spike 率,再將嗰 10 段間隔嘅發射頻率合埋計個平均值出嚟。即係話,發射頻率()等如[27]:
當中 係 咁長嘅時間之內嘅 spike 數量。
喺計發射頻率嗰陣,「用嘅時間間隔要有幾長」都係一個重大嘅問題。用神經發放率計發射頻率會睇唔到一次試驗當中發射頻率嘅變化:呢種直接計法喺研究緊嗰個外界刺激好慢或者好恆定嗰陣最啱用,因為喺呢種情況下(至少理論上)可以預期發射頻率唔會點變;不過,現實世界有好多外界刺激都係會快速噉轉變嘅,而且就算係(例如)喺望住一幅靜止唔郁嘅圖嗰陣,個人對眼都會郁嚟郁去-視網膜上所得到嘅影像會每隔幾百毫秒就變一次,而喺呢種情況下,發射頻率就理論上有可能會喺一次試驗當中變化,神經發放率計法就會搞到研究者睇唔到呢啲變化。總體嚟講,神經發放率有用,但都有缺憾[27]。
時相關射率
[編輯]時相關射率(time-dependent firing rate)係發射頻率嘅另外一種計法,涉及考慮多次重複實驗嘅頻率,定義係喺時間點 同時間點 之間嗰段短時間內嘅平均 spike 數量(),做法係要攞咗「每次試驗嘅 spike 數量」()先,而每次試驗嘅時間長度係 ,再將得出嘅數值除以 ,用公式表達嘅話就係[27]:
喺一個用呢種計法嘅神經編碼實驗裏面,研究者會量度一粒(或者一柞)受緊某啲刺激嘅神經細胞;同一樣嘅刺激會重複好多次,而個研究者會監察住啲神經細胞喺每個時間點入面嘅電壓, 嘅值要有返咁上下大(通常係幾毫秒),先可以確保每段 入面都會有最少一兩個 spike,令到個研究者能夠可靠噉估計 嘅值。攞到嗮呢啲數據之後,就可以跟上面嗰條式,計返個 值出嚟。 話到俾研究者聽喺每段 之間通常有幾多 spike,反映到研究緊嗰粒或者嗰柞神經細胞有幾活躍[27][28]。
時相關射率無論係研究靜止定係變緊嘅外界刺激都有用,但呢種計法又係有佢嘅局限:喺現實情況入面,一粒神經細胞有可能會喺每次受刺激嗰陣運算吓喺下一段時間入面要射幾多 spike,但佢冇乜可能會吓吓都等啲刺激重複幾次先至做反應,所以神經系統冇可能真係靠睇時相關射率傳達資訊嘅。唯有喺研究緊一大柞一齊受刺激嘅神經細胞嗰陣,時相關射率先至可以話係大致上準確噉描述現實嘅[27]。

簡史
[編輯]頻率編碼呢個諗頭係由英格蘭生理學家艾加·堄著仁(Edgar Adrian)同瑞典神經科學家榮格·佐塔蒙(Yngve Zotterman)喺 1926 年嗰時首次提出嘅。佢哋做咗個實驗,將幾嚿唔同重量嘅物件吊喺一條肌肉上面,發現到嚿物件嘅重量愈重,佢哋由條感覺神經嗰度量度到嘅 spike 嘅數量就愈多;由呢個實驗嗰度,埃德里安同佐塔蒙提出話每一個動作電位係一個單位嘅事件,而神經細胞之間嘅溝通唔係建基喺每一件事件嘅大細上嘅,呢啲事件嘅頻率先至係神經細胞之間資訊內容嘅所在-就有咗頻率編碼嘅諗頭[29]。
埃德里安同佐塔蒙嗰份研究相當有影響力,再加上 spike 嘅發射頻率又相對易量度,所以喺嗰份研究打後嘅嗰幾廿年,神經科學家之間有咗股研究發射頻率嘅風潮,甚至有啲神經科學家仲主張話神經編碼研究淨係需要知道發射頻率就得。但廿一世紀初開始嘅研究經已表明,有好多發射頻率以外嘅因素都會反映到神經脈衝資訊嘅內容[24],而且呢種情況喺某啲特定類型嘅神經細胞入面零舍會發生。所以廿一世紀嘅神經編碼研究一般都唔會再齋睇發射頻率[30][31]。
時間編碼
[編輯]
時間編碼(temporal coding)係另一種編碼方案,指啲 spike 帶嘅資訊有至少一部份係透過 spike 嘅(相對)出現時間[2]、發射頻率嘅上落、或者係 spike 之間嘅時間間隔(inter-spike intervals,ISI)等嘅時間性特徵嚟表達嘅,即係話(1 代表有 spike,0 代表冇 spike) 000111000111 同 001100110011 呢兩串時間序列-兩者都係喺 12 毫秒內有 6 個 spike-表達嘅資訊可以唔同[32][33]。
事實係有多份研究都廿世紀尾至廿一世紀初嘅研究表明,時間編碼嘅諗頭真係掂。首先,神經訊號嘅時間解像度去到成以毫秒計,噉表示 spike 嘅出現時間有可能做到有效噉表示複雜嘅資訊[2][34]。另一方面,仲有進一步研究發現以下呢兩點:
- 發射頻率極之相似嘅 spike 列可以傳達到唔同資訊,表示咗 spike 列當中嘅資訊唔係淨係由發射頻率表達嘅[35],而
- 對靈長目嘅大腦皮層前部份嘅研究又顯示,就連得嗰幾毫秒咁長嘅極短暫 spike 規律有陣時都能夠預測動物處理資訊嘅行為,呢啲極短暫嘅 spike 規律冇乜可能係靠頻率嚟傳資訊嘅[35]。
一般認為上述兩個事實表明咗,「spike 嘅出現時間」等嘅特徵的確會幫手傳達資訊[36]。
發射相位編碼
[編輯]發射相位編碼(phase-of-firing code)係一種特殊嘅神經編碼方案,指將粒神經細胞傳緊嘅資訊內容以 spike 喺神經振盪(neural oscillation)當中嘅相位(phase)嚟傳達:神經振盪,俗稱「腦電波」,指中央神經系統入面神經活動週期性噉上上落落嘅節奏,而一個 spike 嘅相位指嗰個 spike 喺一個活動週期入面嘅位置,用角度表示-即係 0 度表示個 spike 喺個週期嘅起始點出現、180 度表示個 spike 喺個週期嘅正中間點出現... 如此類推[37][38]。
事實表明,如果神經網絡嘅活動嘅上落嘅頻率低嘅話,用相位嚟做嘅編碼當中通常「相位」淨係需要有 4 個離散(discrete)嘅數值,就經已夠攞嚟代表嗮呢種訊號要代表嘅資訊內容-係一種相當有效率嘅編碼方法,而且高頻嘅神經振盪都可以用發射相位編碼[39]。而實證嘅研究顯示,喺大腦皮層嘅某啲感覺區域當中,「spike 出現嘅時間」相對於「神經網絡嘅活動嘅上落嘅相位」嘅相位數值的確能夠預測外界刺激嘅特性[37]。
感官時間編碼
[編輯]快度感知
[編輯]一般認為,時間編碼解釋得到點解動物識得處理一啲嚟得快去得快嘅資訊:當要應對一啲好短暫嘅外界刺激(例如見到一件以高速度郁動嘅物體)嗰陣,一粒神經細胞能夠及時產生得到多過一個 spike-而得嗰一個 spike 就表示,呢啲資訊冇可能係齋靠頻率編碼嚟傳遞嘅;之但係事實又表明,動物真係有能對呢啲短暫嘅外界刺激俾反應[40]。
「時間編碼幫到手處理極之短暫嘅資訊」亦都俾人指係解釋到感官點樣用聲定位:有聽覺嘅動物能夠或多或少噉靠聽周圍嘅聲嚟感知外界物件嘅位置;蝙蝠同海豚等嘅某幾種動物就零舍擅長做呢樣嘢-佢哋勁到可以靠自己射啲聲波出去再聽啲聲點樣彈返嚟,做到高度準確噉判斷周圍嘅每嚿物體離自己有幾遠,並且靠噉嚟捕殺以高速度飛緊或者游緊嘅獵物,呢種能力就係所謂嘅反響定位(echolocation);呢種能力要求個神經系統曉喺以毫秒計嘅極短時間間隔內攞大量嘅聽覺資訊,例如係「感官每個位聽到嘅聲係喺邊個時間點聽到嘅」;電腦模擬等研究嘅結果顯示,如果呢啲資訊要純粹靠(可以慢得好交關)嘅發射頻率嚟傳達嘅話,噉一粒等緊聽覺資訊嘅神經細胞可以要等成超過 1 秒先能夠區分唔同嘅聽覺資訊-冇可能會做得到喺實驗觀察到嘅嗰種超快速聲音定位[40]。

唔同感官
[編輯]事實表明,唔同感官都有用到時間編碼:
- 視覺編碼:為咗解釋神經細胞點樣快速噉傳遞視覺資訊,有科學家主張視網膜(retina;眼珠入面一浸能夠感光嘅膜)會將視覺資訊放喺「刺激出現嘅時間」同「第一個 spike 出現」之間嘅時間間隔當中[41],而呢個數就係所謂嘅到第一個 spike 嘅潛伏期(latency to first spike)。雖然呢種傳資訊方法比較易受雜音干擾,但就相當有用-有神經科學家做過研究,發現喺獼猴嘅視覺皮層(visual cortex)入面,到第一個 spike 嘅潛伏期提供到嘅資訊多過 ISI 所能夠提供嘅(簡單講,即係「到第一個 spike 嘅潛伏期」更加能夠預測感知到嘅資訊),而且「用到第一個 spike 嘅潛伏期傳達資訊」嘅現象喺動物嘅聽覺同觸覺系統入面都有出現[42]。
- 味覺編碼:哺乳類動物嘅味覺系統所受嘅刺激(掂到嘅嘢嘅化學成份)可以分做幾個簡單嘅大類-甜、酸、苦同鹹呀噉,而且啲哺乳類對唔同嘅味道又會有好明顯唔同嘅反應,所以哺乳類味覺系統對於研究時間編碼嚟講好有用;用時間性特徵嚟傳達嘅資訊會幫到隻動物分辨同一種味道嘅唔同分子,而每類味道都會產生一個獨特規律嘅 spike 反;一個味道分子所產生嘅 spike 反應嘅時間特徵可以幫到研究者預測道隻佢係乜味-頻率會俾到有關「嗰個味道屬邊個類」嘅資訊,而時間性特徵就會俾到再仔細啲嘅資訊,幫到個研究者分(例如)唔同種嘅苦味分子。即係話味覺系統會同時用時間編碼同頻率編碼嚟傳資訊,而兩種編碼方法傳嘅資訊嘅內容唔同[43][44]。
- 嗅覺編碼:研究顯示,有多種動物嘅嗅覺系統都有用到時間編碼。喺家鼠嘅嗅球當中,到第一個 spike 嘅潛伏期會傳達到有關氣味嘅資訊;有研究者指,同頻率編碼比起上嚟,時間編碼能夠更加快噉傳資訊,令到隻老鼠行動起上嚟更加快,有助佢生存[45]。同一道理,實驗顯示,兔仔同蝗蟲嘅嗅覺感官系統都會用時間編碼嚟到傳達「聞到乜嘢味」呢個資訊俾個腦知[2]。
時間編碼嘅進化
[編輯]由進化論嘅角度睇,時間編碼仲能夠解釋到點解神經細胞嘅發射頻率可以有高頻率嘅變化(變得好頻繁):如果神經系統淨係用頻率編碼嚟傳資訊嘅話,噉比較恆定不變嘅發射頻率會喺進化上有利(有助隻動物生存同繁殖),所以神經細胞理應會進化成採用呢種比較恆定嘅編碼方式;但實際上,發射頻率仲係有好多上上落落,所以「神經系統純粹用頻率編碼嚟傳資訊」呢個假說會做一個同觀察到嘅事實唔夾嘅預測[46];相比之下,有咗時間編碼呢個概念嘅話就解得通-發射頻率嘅嗰啲變化唔係雜音,而係真係對神經細胞傳資訊有用嘅嘢,所以先會通過物競天擇嘅考驗留存到落嚟[40]。
總體編碼
[編輯]

頻率編碼同時間編碼都係用個別一粒粒神經細胞射嘅 spike 嘅規律嚟傳達資訊嘅,而總體編碼(population coding)呢個方案就係指用一大柞神經細胞整體嘅活動規律嚟傳達資訊。喺總體編碼當中,每一粒神經細胞都有獨特嘅一套反應規律,都會各自對唔同嘅輸入作出唔同嘅反應,而且一柞神經細胞當中可能有啲仲會冇反應;而呢柞神經細胞嘅反應夾埋一齊嘅規律會幫到睇嗰個人預測呢啲神經細胞受到咗啲乜嘢輸入[47][48]。
數學性啲噉講嘅話,總體編碼可以用總體向量(population vector)嘅概念嚟想像[49]:
當中 係一個腦區第 粒神經細胞嘅活動,而 表示神經細胞 偏好嗰個輸入[註 4]。舉個簡單例子說明,用 -1 代表「左」、+1 代表「右」,而家吓個腦望到有件物體向左手邊郁緊,噉偏好向左手邊郁嘅嘢嗰啲神經細胞會有好勁嘅活動(佢哋 數值高),而唔偏好向左手邊郁嘅嘢嗰啲細胞嘅活動會相對弱(佢哋 數值低),於是用上面條式計到出嚟嗰個 數值會偏向左手邊(-1)-呢個腦區成功噉傳達咗有關件物體嘅移動方向嘅資訊[49]。
實驗性嘅研究顯示咗,總體編碼嘅做法喺個腦嘅感覺同運動區域嗰度都有出現。例如係幫手管視覺嘅中顳葉(medial temporal lobe,簡稱「MT」)噉,嗰度啲神經細胞會對物件郁嘅方向有反應,當個人望到一嚿向住某個特定方向郁緊嘅物體嗰陣,MT 嘅神經細胞整體上會以一個帶有雜音同常態分佈嘅規律射 spike,而嚿物體郁嘅方向會決定呢個規律嘅型態,而個別神經細胞嘅射 spike 規律就算同 MT 個整體唔同,都唔會影響最後成個 MT 得出嘅訊號[50];運動控制方面嘅例子就有主要運動皮層(primary motor cortex),有神經科學家做咗研究,訓練馬騮去操作一碌遊戲搖桿,要啲馬騮將支搖桿移向一嚿發光嘅嘢嗰度;結果發現,一粒個別嘅神經細胞會對多個方向起反應,但每粒都有一個特定嘅方向,喺隻馬騮要將支搖桿移向嗰個方向嗰陣射 spike 射得零舍快,所以呢柞神經細胞嘅整體射 spike 規律(邊個方向對應嗰啲細胞射 spike 射得最快)會反映到隻馬騮喺度將支搖桿移緊向邊個方向[51]。
- 理論優點
一般認為,總體編碼有好多好處:
- 由理論角度嚟睇,總體編碼係一個喺數學上有咗精細解法嘅問題:總體編碼相對簡單,要用運算模型模擬佢好易,唔似得頻率編碼或者時間編碼噉,吓吓都講個別神經細胞嘅活動-因為一個腦區閒閒地都有成幾億粒神經細胞,所以做起電腦模擬上嚟好多時都會好撈絞-電腦模擬頻率編碼或者時間編碼往往要將嗰成幾億粒細胞逐粒逐粒模擬嗮,而電腦模擬總體編碼好多時都淨係使考慮「估計有幾多百分比嘅細胞以呢種規律射緊 spike」等嘅問題,相比之下就簡單好多[52];
- 總體編碼仲能夠減少不確定性(uncertainty)-一粒個別嘅神經細胞容易俾雜音影響,搞到佢訊號走樣;但(例如)「1,000 粒神經細胞大部份都出錯」嘅機會率就低過「一粒神經細胞出錯」嘅機會率好多,所以用大柞神經細胞嚟傳資訊理論上係會準啲精確啲[52][53]。
位置編碼
[編輯]位置編碼(position coding)係總體編碼嘅一種。喺一個總體編碼入面,「所受嘅外界刺激嘅特徵」同「神經細胞嘅整體活動」好多時會形成高斯函數(Gaussian function),個函數如下:
當中 係一個刺激嘅特徵(例如一束光嘅亮度),當中 、 同 係描述粒細胞或者柞細胞嘅參數[54]。如果一粒神經細胞跟高斯函數( 係活動嘅強度, 係刺激特徵)而一個外界刺激嘅數值同特定平均值()一樣,,就會引致粒細胞有最強嘅活動,而當一個外界刺激嘅數值偏離 , 數值上升,就會引致 數值下降-即係話一粒跟高斯函數嘅神經細胞會對所有刺激有反應,但對佢偏好嗰個特定刺激數值會有最強嘅反應[55]。
想像有一柞神經細胞,每粒細胞嘅 值都唔同,但多數嘅細胞 值都傾向於某個特定數值(, ≈ 啲細胞嘅平均 )。當有一個數值接近 嘅刺激出現,就會引致多數細胞有強嘅活動,而當有一個數值離 遠嘅刺激出現,柞細胞都會有活動,但整體活動就弱。所以如果用 X 軸代表「所受嘅外界刺激嘅特徵」,Y 軸代表「神經細胞嘅整體活動」,噉畫出嚟嗰條線會好似個鐘噉嘅形(睇埋常態分佈)。呢種編碼主要用嚟傳達一啲連續性變數嘅數值,例如係關節同眼嘅位置、色水或者係聲嘅頻率等等[55]。

相關編碼
[編輯]相關編碼(correlation coding)嘅諗法指出,spike 列之間嘅統計相關(statistical correlation)可以帶有一啲資訊。早期嘅研究認為,spike 列之間嘅相關只係能夠減少-唔能夠提升-兩條 spike 列當中帶嘅資訊[56],但係呢個諗法遲吓就受到新證據挑戰:研究發現,喺某啲情況之下,spike 列同 spike 列之間嘅相關可以提升佢哋所帶嘅資訊嘅內容量[57]。相關亦都可以帶有兩對神經細胞之間嘅個別發射率入面冇嘅資訊,例如有研究指,喺某啲靈長目嘅聽覺皮層嗰度,「聽到純音」會令個腦區有更多彼此之間有高統計相關嘅 spike,但啲神經細胞嘅平均發射率就唔變[58]。
疏編碼
[編輯]疏編碼(sparse coding)係總體編碼嘅一種,指外界刺激嘅每一個部份都由一細組神經細胞嘅強烈活動嚟入碼[59][60]。喺疏編碼當中,每一柞神經細胞會負責處理一啲特定嘅資訊,而呢柞神經細胞會分做幾組,每組專門負責處理嗰啲資訊當中嘅一啲特定部份,所以好多時都淨係得一兩組神經細胞活躍,而淨低嗰啲就冇乜活動-神經活動分佈「稀疏」。廿一世紀初嘅神經科學研究顯示,疏編碼係生物神經系統同傳統電子電腦比起上嚟最唔同嘅地方之一,因為喺生物神經系統當中,同一個資訊可以分佈喺好多粒神經細胞之間,而相比之下,一部電腦運算緊嘅每一份資訊都淨係會由一至兩粒處理器集中負責嗮[61][62]。
有人指,疏編碼表示資訊嘅方法有啲似人類語言:當一柞神經細胞收到一柞量可以好大嘅輸入嗰陣,用疏編碼嘅演算法會嘗試搵一細組能夠用嚟表示呢啲輸入嘅神經活動規律,而當呢啲活動規律以啱嘅比例或者次序結合埋一齊嗰陣,就會產生原本嗰個輸入,而呢啲活動規律就係呢啲輸入嘅疏編碼-似十足語言噉;舉個例說明,粵文入面會出現嘅句子可以有好多好多唔同嘅款,但係呢啲款冚唪唥都可以用相對少量嘅符號-即係漢字、粵字同標點符號呀噉-以某啲特定嘅次序排列結合埋一齊嚟到傳達[61]。
線性生成模型
[編輯]多數疏編碼嘅模型都係建基於線性生成模型(linear generative model)嘅,喺呢啲模型當中,符號會以線性嘅型式結合埋一齊去產生收到嗰個輸入[63]。正式啲嚟講,線性生成模型定義如下:如果家吓個輸入用一個有 咁多個維嘅向量代表,疏編碼嘅目的係要搵出 個 維嘅基向量(basis vector)嚟代表 粒神經細胞,以及每個呢啲基向量都要有一個 維、疏嘅(指啲元素多數係 0 嘅)向量嚟代表呢粒神經細胞有幾影響到個輸出,令到以下呢條式就會俾到個啱嘅輸出[64]:
- ,
當中:
- ,係有 個維度,代表輸入嘅向量;
- ,有 個分別代表 粒神經細胞嘅向量;
- ,有 n 個權重,第 個權重代表第 粒神經細胞有幾影響到個輸出。
完整度
[編輯]另一種將線性生成模型演算法整出嚟嘅編碼分類嘅方法,係睇佢係啱啱好完整(critically complete)定係完整過龍(overcomplete)。如果神經細胞嘅數量 啱啱好等如輸入向量嘅維度 ,噉呢個編碼就算係「啱啱好完整」,喺呢個情況下,輸入向量嘅改變可以引致神經細胞向量嗰啲系數突然間改變得好犀利,而呢個編碼唔會曉平滑噉處理輸入嗰度嘅雜音或者輸入嘅縮放;但如果神經細胞嘅數量 大過輸入向量嘅維度 ,噉呢個編碼就謂之「完整過龍」,能夠平滑噉處理輸入嗰度嘅雜音或者輸入嘅縮放[65]。完整過龍嘅編碼好好用:據估計,人類嘅主要視覺皮層(primary visual cortex)完整過龍咗成差唔多 500 倍,一個 14 x 14 (196 個維度)嘅輸入會有成 100,000 粒神經細胞負責處理運算[63]。
疏編碼嘅證據
[編輯]喺廿一世紀初,有神經科學研究者指,疏編碼或者可以解釋到神經系統普遍係點樣做記憶功能嘅:為咗要適應四圍嘅環境,一隻動物實要去學識分辨邊啲外界刺激對自己有益(睇埋行為強化),邊啲對自己有害(睇埋行為懲罰),邊啲同自己嘅生存唔啦更;呢種認知作業要求隻動物識得用記憶功能,將自己經歷過嘅嘢記低,而喺呢個記憶過程入面,是但攞一個外界刺激,一個總體嘅神經細胞當中得一少部份(負責處理呢個刺激嗰啲細胞)會對呢個刺激有反應,而每粒神經細胞都淨係會對少數幾個外界刺激有反應-就好似疏編碼描述嘅過程噉樣[66]。喺實驗上,科學家經已發現咗有多個神經系統都會用疏編碼代表視覺[67]、聽覺[68]、觸覺[69]同嗅覺[70]等方面嘅感覺資訊。
應用
[編輯]配合光遺傳學
[編輯]時間編碼可以配合光遺傳學(optogenetics),達致自由噉控制一隻動物嘅神經系統活動:光遺傳學呢種技術涉及控制一隻動物嘅基因,將佢啲神經細胞變成會對光有反應,而跟手科學家就可以用光控制呢啲受改造嘅神經細胞,有得(例如)控制個別神經細胞產生乜嘢同點樣嘅 spike,而且解像度高到睇到個別單一細胞嘅程度[72];舉個例說明,想像而家有粒受改造嘅老鼠神經細胞,佢會對藍色光有反應,每次受藍色光刺激嗰陣都會產生一個 spike;而當粒細胞感應唔到藍光嗰陣時,粒細胞就唔會再產生 spike,即係話研究者會能夠靠藍光訊號嚟控制神經細胞產生 spike [73]。然後神經科學家就能夠透過操弄隻老鼠嘅神經細胞嘅 spike 發射,睇吓 spike 嘅發射時間傳達到乜嘢資訊(例如係睇吓 ISI 嘅變化會點樣影響老鼠嘅肌肉活動)[74]。
除咗科研上有用之外,「用光遺傳學控制 spike 嘅時間」呢種技術仲俾人指係有可能幫到手醫一啲同 spike 異常有關嘅神經或者心理疾病[74]。透過控制 spike 嘅產生,神經內科醫生可以特登提高或者壓抑個腦某啲區域嘅活動,等個腦嘅活動變成正常嘅樣-而好多嘅神經或者心理疾病,包括咗抑鬱症(major depression)、精神分裂(schizophrenia)同柏金遜症(Parkinson's disease)呀噉,都係源於個腦某啲區域嘅活動異常嘅。而且有研究顯示,對 spike 與 spike 之間嘅時間間隔嘅操作喺控制起腦活動嗰陣,可能仲有效過藥[73]。
智能放大
[編輯]智能放大(intelligence amplification,簡稱「IA」)係認知科學等領域上嘅一個概念,泛指一啲運用資訊科技協助提升人類智能嘅做法[75]。舉個簡單例子說明,想像有個工程師創造出一個裝置,能夠將一個人腦接駁落去一部能夠連接互聯網嘅電腦嗰度,令個人腦同部電腦有得互相直接傳達資訊,而個人腦又可以俾指令(即係齋靠諗嚟指揮一部電腦),要部電腦用搜尋器耖資料,然後再將啲資料直接傳返去個人腦嗰度-就會做到「用人工裝置提升一個人嘅智能」嘅效果。呢個過程當中要用到神經編碼嘅知識,因為上述嘅裝置最少需要做到兩樣嘢-將人腦傳嘅訊號解讀成電腦睇得明嘅訊號(神經解碼),然後又要將電腦俾返嚟嘅輸出變做個人腦睇得明嘅訊號(神經入碼)[76][77]。

註釋
[編輯]睇埋
[編輯]相關技術
[編輯]文獻
[編輯]- Borst, A., & Theunissen, F. E. (1999). Information theory and neural coding (PDF). Nature neuroscience, 2(11), 947-957.
- Dayan P & Abbott LF. Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems. Cambridge, Massachusetts: The MIT Press; 2001. ISBN 0-262-04199-5
- Rieke F, Warland D, de Ruyter van Steveninck R, Bialek W. Spikes: Exploring the Neural Code. Cambridge, Massachusetts: The MIT Press; 1999. ISBN 0-262-68108-0
- Olshausen, B. A., & Field, D. J. (1996). Emergence of simple-cell receptive field properties by learning a sparse code for natural images (PDF). Nature, 381(6583), 607-609.
- Tsien, J. Z. et al. (2014). "On initial Brain Activity Mapping of episodic and semantic memory code in the hippocampus". Neurobiology of Learning and Memory. 105: 200–210.
攷
[編輯]- ↑ "How many neurons make a human brain? Billions fewer than we thought | James Randerson". the Guardian (英文). 2012-02-28. 喺2021-05-25搵到.
- ↑ 2.0 2.1 2.2 2.3 2.4 Thorpe, S. J. (1990). "Spike arrival times: A highly efficient coding scheme for neural networks". In Eckmiller, R.; Hartmann, G.; Hauske, G. Parallel processing in neural systems and computers. North-Holland. pp. 91-94.
- ↑ Brown E. N., Kass R. E., Mitra P. P. (May 2004). "Multiple neural spike train data analysis: state-of-the-art and future challenges". Nat. Neurosci. 7 (5): 456–61.
- ↑ Johnson, K. O. (2000-6). "Neural coding". Neuron. 26 (3): 563–566.
- ↑ 5.0 5.1 The Memory Code. Scientific American.
- ↑ 6.0 6.1 Chen, G.; Wang, L. P.; Tsien, J. Z. (2009). "Neural population-level memory traces in the mouse hippocampus". PLOS ONE. 4(12): e8256.
- ↑ 7.0 7.1 Brain Decoding Project.
- ↑ Zhang, H.; Chen, G.; Kuang, H.; Tsien, J. Z. (Nov 2013). "Mapping and deciphering neural codes of NMDA receptor-dependent fear memory engrams in the hippocampus". PLOS ONE. 8(11): e79454.
- ↑ The Simons Collaboration on the Global Brain.
- ↑ Hodgkin, A. L., & Huxley, A. F. (1952). "A quantitative description of membrane current and its application to conduction and excitation in nerve". The Journal of Physiology. 117 (4): 500–544.
- ↑ Fraser, A.; Frey, A. H. (1968). "Electromagnetic emission at micron wavelengths from active nerves". Biophysical Journal. 8 (6): 731–734.
- ↑ Adrian, E. D. (1964). The Basis of Sensation. Hafner, New York.
- ↑ Roland Priemer (1991). Introductory Signal Processing. World Scientific. p. 1.
- ↑ Amari, SL (2001), Information Geometry on Hierarchy of Probability Distributions, IEEE Transactions on Information Theory 47, 1701-1711.
- ↑ Onken, A.; Grünewälder, S.; Munk, M. H. J.; Obermayer, K. (2009), "Analyzing Short-Term Noise Dependencies of Spike-Counts in Macaque Prefrontal Cortex Using Copulas and the Flashlight Transformation", PLoS Comput Biol, 5 (11): e1000577.
- ↑ Aur, D.; Jog, M.S. (2007). "Reading the Neural Code: What do Spikes Mean for Behavior?". Nature Precedings.
- ↑ Gerstner W., Kreiter A. K., Markram H., Herz A. V. (November 1997). "Neural codes: firing rates and beyond". Proc. Natl. Acad. Sci. U.S.A. 94 (24): 12740–1.
- ↑ Burcas G.T & Albright T.D. Gauging sensory representations in the brain. [1]
- ↑ Aur D., Jog, MS., 2010 Neuroelectrodynamics: Understanding the brain language, IOS Press, 2010.
- ↑ Aur, D.; Connolly, C.I.; Jog, M.S. (2005). "Computing spike directivity with tetrodes". J. Neurosci. 149 (1): 57–63.
- ↑ Aur, D. (2012). "A comparative analysis of integrating visual information in local neuronal ensembles". Journal of Neuroscience Methods. 207 (1): 23–30.
- ↑ Butts, D. A., & Goldman, M. S. (2006). Tuning curves, neuronal variability, and sensory coding. PLoS biology, 4(4), e92.
- ↑ Schneidman, E.; Berry, M. J.; & Segev, R.; Bialek, W. (2006), "Weak Pairwise Correlations Imply Strongly Correlated Network States in a Neural Population", Nature, Nature 440, 1007-1012, 440: 1007–1012.
- ↑ 24.0 24.1 24.2 Stein R. B., Gossen E. R., Jones K. E. (May 2005). "Neuronal variability: noise or part of the signal?". Nat. Rev. Neurosci. 6 (5): 389–97.
- ↑ Gerstner, Wulfram; Kistler, Werner M. (2002). Spiking Neuron Models: Single Neurons, Populations, Plasticity. Cambridge University Press.
- ↑ 26.0 26.1 Kandel, E.; Schwartz, J.; Jessel, T.M. (1991). Principles of Neural Science (3rd ed.). Elsevier.
- ↑ 27.0 27.1 27.2 27.3 27.4 1.5 Rate Codes 互聯網檔案館嘅歸檔,歸檔日期2020年4月13號,..
- ↑ Franklin Bretschneider and Jan R. De Weille (2006). Introduction to Electrophysiological Methods and Instrumentation. Academic Press.
- ↑ Adrian ED, Zotterman Y (1926). "The impulses produced by sensory nerve endings: Part II: The response of a single end organ". J Physiol. 61: 151–171.
- ↑ Forrest MD (2014). "Intracellular Calcium Dynamics Permit a Purkinje Neuron Model to Perform Toggle and Gain Computations Upon its Inputs". Frontiers in Computational Neuroscience. 8: 86.
- ↑ Forrest MD (December 2014). "The sodium-potassium pump is an information processing element in brain computation". Frontiers in Physiology. 5 (472).
- ↑ Dayan, Peter; Abbott, L. F. (2001). Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems. Massachusetts Institute of Technology Press.
- ↑ Kostal L., Lansky P., Rospars J. P. (November 2007). "Neuronal coding and spiking randomness". Eur. J. Neurosci. 26 (10): 2693–701.
- ↑ Butts D. A., Weng C., Jin J., et al. (September 2007). "Temporal precision in the neural code and the timescales of natural vision". Nature. 449 (7158): 92–5.
- ↑ 35.0 35.1 Zador, Stevens, Charles, Anthony. "The enigma of the brain". Current Biology, 1995, Vol 5 No 12.
- ↑ Singh & Levy, "A consensus layer V pyramidal neuron can sustain interpulse-interval coding", PLoS ONE, 2017.
- ↑ 37.0 37.1 Marcelo A. Montemurro, Malte J. Rasch, Yusuke Murayama, Nikos K. Logothetis, Stefano Panzeri, Phase-of-Firing Coding of Natural Visual Stimuli in Primary Visual Cortex, Current Biology, Volume 18, Issue 5, 11 March 2008, Pages 375-380.
- ↑ Fries P., Nikolić D., Singer W. (July 2007). "The gamma cycle". Trends Neurosci. 30 (7): 309–16.
- ↑ Havenith MN, Yu S, Biederlack J, Chen NH, Singer W, Nikolić D (June 2011). "Synchrony makes neurons fire in sequence, and stimulus properties determine who is ahead". J. Neurosci. 31 (23): 8570–84.
- ↑ 40.0 40.1 40.2 Theunissen, F.; Miller, J. P. (1995). "Temporal Encoding in Nervous Systems: A Rigorous Definition". Journal of Computational Neuroscience. 2: 149–162.
- ↑ Gollisch, T.; Meister, M. (22 February 2008). "Rapid Neural Coding in the Retina with Relative Spike Latencies". Science. 319 (5866): 1108–1111.
- ↑ Wainrib, Gilles; Michèle, Thieullen; Khashayar, Pakdaman (7 April 2010). "Intrinsic variability of latency to first-spike". Biological Cybernetics. 103 (1): 43–56.
- ↑ Hallock, Robert M.; Di Lorenzo, Patricia M. (2006). "Temporal coding in the gustatory system". Neuroscience & Biobehavioral Reviews. 30 (8): 1145–1160.
- ↑ Carleton, Alan; Accolla, Riccardo; Simon, Sidney A. (2010). "Coding in the mammalian gustatory system". Trends in Neurosciences. 33 (7): 326–334.
- ↑ Wilson, Rachel I. (2008). "Neural and behavioral mechanisms of olfactory perception". Current Opinion in Neurobiology. 18 (4): 408–412.
- ↑ J. Leo van Hemmen, T. J. Sejnowski. 23 Problems in Systems Neuroscience. Oxford Univ. Press, 2006. p.143-158.
- ↑ Dayan P. & Abbott L. F. Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems. Cambridge, Massachusetts: The MIT Press; 2001.
- ↑ Rieke F., Warland D., de Ruyter van Steveninck R., Bialek W. Spikes: Exploring the Neural Code. Cambridge, Massachusetts: The MIT Press; 1999.
- ↑ 49.0 49.1 Mahan M.Y., Georgopoulos A.P. (2014) Neuronal Population Vector. In: Jaeger D., Jung R. (eds) Encyclopedia of Computational Neuroscience. Springer, New York, NY.
- ↑ Maunsell J. H., Van Essen D. C. (May 1983). "Functional properties of neurons in middle temporal visual area of the macaque monkey. I. Selectivity for stimulus direction, speed, and orientation". J. Neurophysiol. 49 (5): 1127–47.
- ↑ Science. 1986 Sep 26;233(4771): 1416-9.
- ↑ 52.0 52.1 Wu S., Amari S., Nakahara H. (May 2002). "Population coding and decoding in a neural field: a computational study". Neural Comput. 14 (5): 999–1026.
- ↑ Hubel DH, Wiesel TN (October 1959). "Receptive fields of single neurones in the cat's striate cortex". J. Physiol. 148 (3): 574–91.
- ↑ Squires, G. L. (2001-08-30). Practical Physics (4 ed.). Cambridge University Press.
- ↑ 55.0 55.1 Mathis A., Herz A. V., Stemmler M. B. (July 2012). "Resolution of nested neuronal representations can be exponential in the number of neurons". Phys. Rev. Lett. 109 (1): 018103.
- ↑ Johnson, K. O. (Jun 1980). "Sensory discrimination: neural processes preceding discrimination decision". J Neurophysiol. 43 (6): 1793–815.
- ↑ Panzeri; Schultz; Treves; Rolls (1999). "Correlations and the encoding of information in the nervous system". Proc Biol Sci. 266 (1423): 1001–12.
- ↑ Merzenich, M. M. (Jun 1996). "Primary cortical representation of sounds by the coordination of action-potential timing". Nature. 381 (6583): 610–3.
- ↑ Olshausen, Bruno A.; Field, David J. (1996). "Emergence of simple-cell receptive field properties by learning a sparse code for natural images". Nature. 381 (6583): 607–609.
- ↑ Gupta, N.; Stopfer, M. (6 October 2014). "A temporal channel for information in sparse sensory coding". Current Biology. 24 (19): 2247–56.
- ↑ 61.0 61.1 What is population coding? Describe the population coding model proposed by Georgopoulos in the 1980s for M1 control of arm direction. Foundational Neuroscience Questions.
- ↑ P. Földiák, 2002, Sparse coding in the primate cortex , in The Handbook of Brain Theory and Neural Networks, Second Edition, pp 1064-1068, ed. Michael A. Arbib, MIT Press.
- ↑ 63.0 63.1 Rehn, Martin; Sommer, Friedrich T. (2007). "A network that uses few active neurones to code visual input predicts the diverse shapes of cortical receptive fields". Journal of Computational Neuroscience. 22: 135–146.
- ↑ Lee, Honglak; Battle, Alexis; Raina, Rajat; Ng, Andrew Y. (2006). "Efficient sparse coding algorithms". Advances in Neural Information Processing Systems.
- ↑ Olshausen, Bruno A.; Field, David J. (1997). "Sparse Coding with an Overcomplete Basis Set: A Strategy Employed by V1?". Vision Research. 37 (23): 3311–3325.
- ↑ Nadal, J. P. (1991). Associative memory: on the (puzzling) sparse coding limit. Journal of Physics A: Mathematical and General, 24(5), 1093.
- ↑ Vinje, W. E.; Gallant, J. L. (2000). "Sparse coding and decorrelation in primary visual cortex during natural vision". Science. 287: 1273–1276.
- ↑ Hromádka, T.; Deweese, M. R.; Zador, A. M. (2008). "Sparse representation of sounds in the unanesthetized auditory cortex". PLoS Biol. 6: e16.
- ↑ Crochet, S.; Poulet, J. F. A.; Kremer, Y.; Petersen, C. C. H. (2011). "Synaptic mechanisms underlying sparse coding of active touch". Neuron. 69: 1160–1175.
- ↑ Ito, I; Ong, RCY; Raman, B; Stopfer, M (2008). "Sparse odor representation and olfactory learning". Nat Neurosci. 11: 1177–1184.
- ↑ Liu Y., LeBeouf B., Guo X., Correa P. A., Gualberto D. G., Lints R., Garcia L. R. (March 2011). Goodman M. B. (ed.). "A cholinergic-regulated circuit coordinates the maintenance and bi-stable states of a sensory-motor behavior during Caenorhabditis elegans male copulation". PLOS Genetics. 7 (3): e1001326.
- ↑ Mancuso, J. J.; Kim, J.; Lee, S.; Tsuda, S.; Chow, N. B. H.; Augustine, G. J. (2010). "Optogenetic probing of functional brain circuitry". Experimental Physiology. 96 (1): 26–33.
- ↑ 73.0 73.1 Karl Diesseroth, Lecture. "Personal Growth Series: Karl Diesseroth on Cracking the Neural Code." Google Tech Talks. November 21, 2008.
- ↑ 74.0 74.1 Han X., Qian X., Stern P., Chuong A. S., Boyden E. S. Informational lesions: optical perturbations of spike timing and neural synchrony via microbial opsin gene fusions. Cambridge, Massachusetts: MIT Media Lad, 2009.
- ↑ Carpenter, Carl (2019-11-06). "IA > AI". Medium (英文). 喺2021-05-25搵到.
- ↑ Elon Musk's next reported project is a company dedicated to linking human brains to computers 互聯網檔案館嘅歸檔,歸檔日期2020年4月5號,.. Business Insider Australia.
- ↑ The Surgeon Who Wants to Connect You to the Internet with a Brain Implant. MIT Technology Review.
拎
[編輯]- Földiák P, Endres D, Sparse coding, Scholarpedia, 3(1): 2984, 2008.
