Lípido
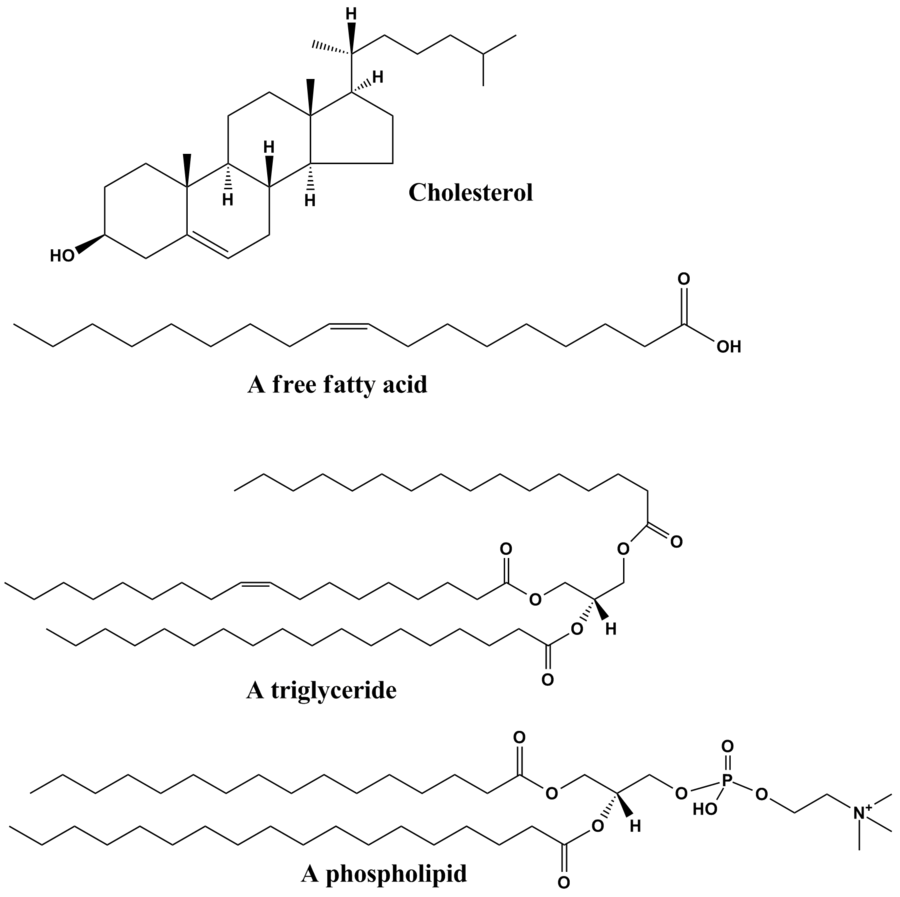

Os lípidos son un amplo grupo de compostos químicos orgánicos naturais, que son uns dos principais compoñentes dos seres vivos, formados principalmente por carbono, hidróxeno e osíxeno, aínda que tamén poden conter fósforo, nitróxeno e xofre, entre os que se inclúen as graxas, ceras, esterois, vitaminas liposolubles (como as vitaminas A, D, E, e K), fosfolípidos e outros. Algúns lípidos son moléculas liñais ou curvadas e outros son compostos cíclicos. Poden definirse a grandes trazos como moléculas relativamente pequenas hidrófobas ou anfipáticas. As hidrófobas poden disolverse en solventes apolares. As anfipáticas teñen unha parte polar e outra non polar, e poden formar en disolución en medio acuoso estruturas como vesículas, liposomas, ou membranas. Entre as principais funcións biolóxicas dos lípidos están o almacenamento de enerxía, ser moléculas estruturais nas membranas e intervir na sinalización celular.[4][5] Os lípidos teñen aplicacións en cosmética e na industria alimentaria e a nanotecnoloxía.[6]
Os lípidos biolóxicos orixínanse enteiramente ou en parte a partir de dous tipos de subunidades bioquímicas, que son: grupos cetoacil e isopreno.[4] Tendo en conta este enfoque, os lípidos poden dividirse en varios tipos: acilos graxos (ácidos graxos), glicerolípidos, fosfoglicéridos (ou glicerofosfolípidos), esfingolípidos, sacarolípidos, e policétidos, derivados da condensación de subunidades cetoacil; e lípidos esterois (esterois e esteroides) e lípidos prenólicos (isoprenoides), derivados da condensación de subunidades de isopreno.[4]
Aínda que o termo lípido se usa ás veces como sinónimo de graxa, en realidade ten un significado máis amplo, xa que, propiamente falando, as graxas serían só os triglicéridos. Moitos lípidos conteñen ácidos graxos como compoñente principal (triglicéridos, fosfolípidos). Os ácidos graxos presentan unha reacción característica chamada saponificación cando os facemos reaccionar quimicamente con álcalis na que se orixinan xabóns. Os lípidos con ácidos graxos que presentan esta reacción de saponificación denomínanse saponificables.[7]
Os humanos e outros mamíferos teñen diversas rutas metabólicas para sintetizar a maioría dos seus lípidos ou para degradalos, pero algúns lípidos chamados ácidos graxos esenciais non poden ser sintetizados a partir de precursores simples no organismo, polo que deben tomarse na dieta.
Tipos de lípidos
[editar | editar a fonte]En 2005 o Comité Internacional de Clasificación e Nomenclatura dos Lípidos (ILCNC) publicou un “Sistema de Clasificación Comprensivo para os Lípidos”, que os divide en dous grandes grupos segundo as subunidades que os forman e en oito categorías, que son:[4]
- Lípidos orixinados enteiramente ou en parte por condensacións baseadas en carbanións de tioésteres. Entre eles están os acilo graxos, glicerolípidos, glicerofosfolípidos, esfingolípidos, sacarolípidos e policétidos.
- Lípidos orixinados por condensacións baseadas en carbocatións de unidades de isopreno. Comprende dous grupos: lípidos esterois e lípidos prenólicos.
A continuación a clasificación en detalle cos nomes entre parénteses dos grupos principais que comprende ou sinónimos tradicionalmente usados.
Acilos graxos (ácidos graxos)
[editar | editar a fonte]- Artigo principal: Ácido graxo.
Os ácidos graxos, ou os residuos de ácidos graxos que forman parte dos lípidos, son un grupo moi diverso de moléculas sintetizadas a partir dunha molécula inicial de acetil-CoA, que se alonga con grupos de dous carbonos procedentes de moléculas de malonil-CoA ou metilmalonil-CoA mediante un proceso chamado síntese de ácidos graxos.[8][9] Están formados por unha cadea hidrocarbonada que termina nun grupo ácido carboxílico; esta disposición confírelle á molécula un carácter anfipático, xa que ten un extremo polar ou hidrófilo e outro non polar ou hidrófobo, que é insoluble en auga. Os ácidos graxos son un dos constituíntes máis importantes dos lípidos biolóxicos, e forman parte de moitos lípidos complexos.[10] A cadea carbonada, que xeralmente ten entre 4 e 24 carbonos,[11] pode ser saturada (todos os enlaces entre carbonos simples) ou insaturada (algún enlace dobre ou triplo), e pode levar unidos grupos funcionais que conteñen oxíxeno, nitróxeno, xofre, e halóxenos. Os enlaces dobres da cadea poden ser de tipo cis ou trans, o que a fecta á configuración e propiedades da molécula. Os dobres enlaces cis fan que a cadea do ácido graxo se dobre acusadamente, o que xoga un importante papel na estrutura e función das membranas da célula.[12] A maioría dos ácidos graxos naturais están na configuración cis, aínda que as formas trans existen tamén nalgúns aceites e graxas naturais parcialmente hidroxenados.[13]
Os ácidos graxos máis comúns que forman parte de triglicéridos e fosfolípidos son o ácido palmítico, o esteárico e o oleico. Outros exemplos de ácidos graxos bioloxicamente importantes son os eicosanoides, derivados do ácido araquidónico e o ácido eicosapentaenoico, entre os que se inclúen as prostaglandinas, leucotrienos, e tromboxanos. O ácido docosahexaenoico é tamén importante bioloxicamente, especialmente para a vista.[14][15] Outros lípidos importantes do grupo dos ácidos graxos son os ésteres de ácidos graxos e as amidas de ácidos graxos. Entre os ésteres de ácidos graxos están importantes compostos e intermediarios como as ceras, os derivados tioéster de ácidos graxos e coencima A, o tioéster de ácidos graxos ACP e as ácido graxo carnitinas. Entre as amidas de ácidos graxos están as N-aciletanolaminas, como o neurotransmisor cannabinoide anandamida.[16]
Ceras
[editar | editar a fonte]- Artigo principal: Cera.
O grupo tradicional das ceras estaría incluído nesta categoría dos acilos graxos como ésteres de ácidos graxos. Son lípidos formados por un ácido graxo esterificado por un alcohol de cadea longa cun só OH. Son compostos moi hidrófobos con funcións protectoras, que se encontran no recubrimento da epiderme de follas e froitos, ou son compoñentes da graxa dos pelos e plumas ou da cera dos oídos e a cera de abella.
Glicerolípidos
[editar | editar a fonte]- Artigos principais: Glicérido e Triglicérido.
Os glicerolípidos están compostos principalmente por glicerois mono-, di-, e tri-substituídos,[17] e os máis importantes son os triésteres de ácidos graxos e glicerol chamados triglicéridos ou triacilglicerois, que xa non teñen ningún grupo OH libre do glicerol. Nestes compostos, os tres OH do glicerol están esterificados con ácidos graxos. A súa misión é almacenar enerxía, polo que constitúen a maior parte das graxas que se almacenan no organismo (as graxas do tecido adiposo). A hidrólise dos enlaces éster dos triglicéridos e a conseguinte liberación de glicerol e ácidos graxos no tecido adiposo son os pasos iniciais do metabolismo das graxas.[18]
Outras subclases de glicerolípidos son os glicosilglicerois, que presentan un ou máis residuos de azucres unidos directamente ao glicerol por medio de enlace glicosídico. Exemplos deste tipo son os digalactosildiacilglicerois que se encontran nas membranas das plantas[19] e o seminolípido dos espermatozoides de mamífero.[20]
Glicerofosfolípidos (fosfoglicéridos)
[editar | editar a fonte]- Artigo principal: Fosfoglicérido.
Os glicerofosfolípidos ou fosfoglicéridos, son moi comúns nos seres vivos e o compoñente principal da bicapa lipídica das membranas celulares,[21] e están implicados no metabolismo e sinalización celular.[22] Os tecidos nerviosos como o do cerebro conteñen cantidades relativamente grandes de fosfoglicéridos, e as alteracións na súa composición foron relacionadas con varios trastornos neurolóxicos.[23] Un fosfoglicérido típico está formado por un glicerol esterificado con dous ácidos graxos e unha molécula polar unida ao OH que queda libre do glicerol. Os fosfoglicéridos poden subdividirse en distintos tipos, baseándose na natureza do grupo polar de cabeza situado na posición sn-3 do glicerol nos eucariotas e bacterias, ou na posición sn-1 nas arqueas.[24]

Exemplos de fosfoglicerolípidos que se encontran nas membranas biolóxicas son a fosfatidilcolina, fosfatidiletanolamina e fosfatidilserina.
Ademais de serviren como compoñentes principais das membranas da célula e ser sitios de unión para as proteínas extra- e intracelulares, algúns fosfoglicéridos das células eucarióticas, como os fosfatidilinositois e os ácidos fosfatídicos son segundos mensaxeiros ou precursores de segundos mensaxeiros.[25] Aínda que o típico nos fosfoglicéridos é que dous dos grupos OH do glicerol estean esterificados con ácidos graxos de cadea longa, hainos tamén unidos a alquilos e a 1Z-alquenilos (plasmalóxeno), e variantes dialquiléter nas arqueas.[26]
Esfingolípidos
[editar | editar a fonte]- Artigo principal: Esfingolípido.
Os esfingolípidos son unha complicada familia de compostos[27] que comparten unha estrutura común, un esqueleto formado por unha base esfingoide que se sintetiza a partir do aminoácido serina e dun acilo graxo de longa cadea-CoA, que despois é convertido en ceramidas, fosfoesfingolípidos, glicoesfingolípidos e outros compostos. A principal base esfingoide nos mamíferos é a xeralmente denominada esfingosina. As ceramidas (N-acil-base esfingoide) son unha importante subclase de derivados de base esfingoide cun ácido graxo unido por enlace amida. Os ácidos graxos que levan son tipicamente saturados ou monoinsaturados con cadeas de lonxitudes que van de 16 a 26 átomos de carbono.[28]

Os principais esfingolípidos dos mamíferos son as esfingomielinas (ceramida fosfocolinas),[29] mentres que os insectos conteñen principalmente ceramidas fosfoetanolaminas[30] e os fungos teñen fitoceramidas fosfoinositois e grupos de cabeza que conteñen manosa.[31]
Os glicoesfingolípidos son unha familia diversa de moléculas compostas por un ou máis residuos de azucre unidos por enlace glicosídico á base esfingoide. Exemplos destes son os glicoesfingolípidos simples e complexos como os cerebrósidos e gangliósidos.
Lípidos esterois (esterois e esteroides)
[editar | editar a fonte]
Este grupo de lípidos está formado por un esqueleto cíclico básico de catro aneis chamado esterano, e as distintas modificacións e adicións de grupos químicos e cadeas a este esqueleto orixinan os distintos tipos de esteroides. Os esterois, como o colesterol e os seus derivados, teñen un OH no carbono 3 do esqueleto, e son un importante compoñente dos lípidos de membrana,[32] xunto cos fosfoglicéridos e esfingomielinas. Os esteroides, derivados da mesma estrutura de catro aneis, teñen diferentes funcións biolóxicas como hormonas e moléculas de sinalización. Os esteroides de 18 carbonos inclúen o grupo dos estróxenos, e os de 19 carbonos os andróxenos como a testosterona e a androsterona. A subclase de 21 carbonos comprende os proxestáxenos e os glicocorticoides e mineralocorticoides.[33] Os secosteroides comprenden varias formas de vitamina D e caracterízanse por ter aberto o anel B da súa estrutura básica.[34] Outros exemplos de esterois son os ácidos biliares e os seus conxugados,[35] que nos mamíferos son derivados oxidados do colesterol e sintetízanse no fígado. Os equivalentes nas plantas son os fitosterois, como o β-sitosterol, estigmasterol, e brassicasterol; este último composto tamén se usa como biomarcador para o crecemento das algas.[36] O esterol predominante nas membranas celulares dos fungos é o ergosterol.[37]
Lípidos prenólicos (isoprenoides)
[editar | editar a fonte]- Artigo principal: Terpeno.
Os isoprenoides ou terpenos son lípidos prenólicos sintetizados a partir de precursores de cinco unidades de carbono como o pirofosfato de isopentenilo (chamado isopreno activo) e o pirofosfato de dimetilalilo que se producen principalmente pola vía do ácido mevalónico.[38] Os isoprenoides simples (alcohois liñais, difosfatos etc.) fórmanse pola adición sucesiva de unidades de 5 carbonos, e clasifícanse segundo o número desas unidades de terpeno. As estruturas que conteñen máis de 40 carbonos denomínanse politerpenos. Os carotenoides son importantes isoprenoides simples que funcionan como antioxidantes e como precursores da vitamina A.[39] Outra clase de moléculas bioloxicamente importante son as quinonas e hidroquinonas, que conteñen unha cola isoprenoide unida a un núcleo quinonoide que non é de orixe isoprenoide.[40] As vitaminas E e K, e as ubiquinonas, son exemplos desta clase. Os procariotas sintetizan poliprenois (chamados bactoprenois) nos cales a unidade isoprenoide terminal unida a oxíxeno permanece insaturada, mentres que nos poliprenois animais (dolicois) o isoprenoide terminal está reducido.[41]
Sacarolípidos
[editar | editar a fonte]
Os sacarolípidos son compostos nos cales os ácidos graxos están unidos directamente a un esqueleto de azucres, formando estruturas que son compatibles coas bicapas lipídicas. Nos sacarolípidos, un monosacárido substitúe ao glicerol que tería un glicerolípido ou un fosfoglicérido. Os sacarolípidos máis coñecidos son os precursores de glicosamina acilados do Lípido A compoñente dos lipopolisacáridos nas bacterias gramnegativas. As moléculas típicas de lípido A son disacáridos de glicosamina, dos cales se forman derivados con ata sete cadeas de residuos acilo graxos. O mínimo lipopolisacárido que se require para o crecemento de Escherichia coli é Kdo2-Lípido A, un disacárido hexaacilado de glicosamina que é glicosilado con dous residuos de ácido 3-desoxi-D-mano-octulosónico (Kdo).[42]
Policétidos
[editar | editar a fonte]- Artigo principal: Policétido.
Os policétidos sintetízanse por polimerización de subunidades acetil e propionil por encimas típicos e tamén por encimas repetitivos e multimodulares que comparten características de funcionamento coas ácido graxo sintases. Comprende un gran número de metabolitos secundarios e produtos naturais procedentes de fontes animais, vexetais, bacterianas, fúnxicas e mariñas, e presentan unha gran diversidade estrutural.[43][44] Moitos policétidos son moléculas cíclicas con esqueletos que están con frecuencia modificados por glicosilación, metilación, hidroxilación, oxidación, e outros procesos. Moitos axentes antimicrobianos, antiparasíticos e anticancro usados comunmente son policétidos ou derivados de policétidos, como as eritromicinas, tetraciclinas, avermectinas, e epotilonas antitumores.[45]
Funcións biolóxicas
[editar | editar a fonte]As principais funcións dos lípidos son:
- Estrutural. Certos lípidos son a base estrutural das membranas celulares.
- Reserva enerxética. Depósitos de reserva de enerxía nos organismos (triglicéridos).
- Protección. Recubrimentos protectores hidrófobos nas follas, plumas e pelos.
- Illamento térmico, dado que son malos condutores da calor.
- Absorción da enerxía da luz. Algúns son pigmentos fotosintéticos, como os carotenoides e as xantofilas.
- Función hormonal (esteroides) e de sinalización celular.
- Función metabólica e regulatoria: vitaminas A, E, K, D.
Membranas
[editar | editar a fonte]- Artigos principais: Bicapa lipídica e Membrana plasmática.
As células eucarióticas están compartimentalizadas en orgánulos rodeados de membrana que levan a cabo diferentes funcións biolóxicas. Os fosfoglicéridos son o principal compoñente estrutural da bicapa lipídica das membranas celulares, como a membrana plasmática e as membranas intracelulares dos orgánulos. Nas células animais a membrana plasmática separa fisicamente os compoñentes intracelulares do medio extracelular. Os fosfoglicéridos son moléculas anfipáticas, xa que conteñen unha rexión hidrófoba, formada polas colas de ácidos graxos unidos ao glicerol, e outra hidrófila, formada pola cabeza polar onde está o fosfato e unha molécula polar. Isto permite que se orienten nas membranas coa rexión hidrófoba cara ao interior e a hidrófila cara ás superficies da membrana. Ademais dos fosfoglicéridos, nas membranas hai outros compoñentes que non conteñen glicerol, como as esfingomielinas e os esterois (principalmente colesterol nas membranas animais).[46] En plantas e algas, os galactosildiacilglicerois,[47] e o sulfoquinovosildiacilglicerol,[19] o cal carece de grupo fosfato, son tamén compoñentes importantes das membranas dos cloroplastos e orgánulos relacionados e son os lípidos máis abundantes nos tecidos fotosintéticos de plantas superiores e algas e tamén en certas bacterias.

Unha membrana biolóxica é unha forma de bicapa lipídica. A formación de bicapas lipídicas é un proceso favorecido enerxeticamente cando os fosfoglicéridos se encontran nun ambiente acuoso.[48] Isto coñécese como efecto hidrófobo. Nun sistema acuoso, as cabezas polares dos lípidos alíñanse cara ao medio acuoso polar, mentres que as colas hidrófobas minimizan o seu contacto coa auga e tenden a agruparse, formando unha vesícula; e dependendo da concentración do lípido, esta interacción biofísica pode orixinar a formación de micelas, liposomas, bicapas lipídicas ou outras agregacións. O comportamento de fase dos lípidos é unha área de estudo dentro da biofísica.[49][50][51] Cando se disolve unha substancia lipófila ou anfifílica nun medio polar, as moléculas polares (é dicir, a auga nas disolucións acuosas, que son as máis comúns) sitúanse máis ordenadamente arredor da substancia lipófila disolta, xa que a moléculas polares non poden formar enlaces de hidróxeno coas áreas lipófilas da molécula, polo que nun medio acuoso, as moléculas de auga forman unha gaiola de "clatrato" ordenada arredor da molécula lipófila disolvida.[52]
Almacenamento de enerxía
[editar | editar a fonte]Os triglicéridos, almacenados no tecido adiposo, son a principal forma de almacenamento de enerxía en animais e plantas. O adipocito (célula do tecido adiposo) fai unha continua síntese e degradación de triglicéridos nos animais, e a degradación está controlada polo encima lipase, que é sensible ás hormonas.[53] A oxidación completa dos ácidos graxos proporciona un alto contido calórico, de arredor de 9 kcal/g, comparado coas aproximadamente 4 kcal/g que se producen na oxidación de carbohidratos e proteínas. Os paxaros migratorios que teñen que voar longas distancias sen comer utilizan a enerxía almacenada nos triglicéridos para obter a enerxía que necesitan para as súas viaxes. Os animais hibernantes viven das súas graxas.[54]
Sinalización celular
[editar | editar a fonte]- Artigo principal: Sinalización celular.
Algúns lípidos teñen funcións hormonais (esteroides.[55]) ou de sinalización celular. Nos últimos anos recadáronse moitas evidencias de que a sinalización por lípidos é unha parte fundamental da sinalización celular.[56][57] A sinalización por lípidos pode producirse por medio da activación de receptores acoplados á proteína G ou receptores nucleares, e foron identificados lípidos de diversas clases como moléculas de sinalización e segundos mensaxeiros.[58] Entre eles están:
- A esfingosina-1-fosfato, un esfingolípido derivado das ceramida, que é unha potente molécula mensaxeira implicada na regulación da mobilización do calcio,[59] crecemento celular e apoptose.[60]
- O diacilglicerol (DAG) e os fosfatidilinositol fosfatos (PIPs), implicados na activación mediada por calcio da proteína quinase C.[61]
- As prostaglandinas, que son un tipo de eicosanoide derivado de ácido graxo implicado na inflamación e na inmunidade.[62]
- As hormonas esteroides como os estróxenos, testosterona e cortisol, que modulan un gran número de funcións como a reprodución, metabolismo e presión arterial.
- Os oxisterois como o 25-hidroxi-colesterol, que son agonistas do receptor LXR.[63]
- A fosfatidilserina está implicada na marcaxe para a fagocitose de células apoptóticas e fragmentos de células ao quedar exposta na cara extracelular da membrana (normalmente está na cara citosólica).[64][65]
Outras funcións
[editar | editar a fonte]As vitaminas "solubles en graxas" (A, D, E e K), que son lípidos terpénicos, son nutrientes esenciais que se almacenan no fígado e tecidos adiposos e teñen diversas funcións. As acil-carnitinas interveñen no transporte e metabolismo de ácidos graxos cara a dentro e a fóra da mitocondria, onde sufrirán a beta-oxidación.[66] Os poliprenois e os seus derivados fosforilados teñen tamén importantes misións de transporte, como o transporte de oligosacáridos a través das membranas. Os poliprenol fosfato azucres e poliprenol difosfato azucres funcionan en reaccións extra-citoplasmáticas de glicosilación, na biosíntese de polisacáridos extracelular (por exemplo, a polimerización de peptidoglicanos nas bacterias), e na N-glicosilación de proteínas eucarióticas.[67][68] As cardiolipinas son unha subclase de fosfoglicéridos que conteñen catro cadeas acilo e tres grupos glicerol, que son particularmente abiundantes na membrana mitocondrial interna.[69][70][71]; crese que activan encimas implicados na fosforilación oxidativa.[72] Os carotenos e xantofilas son terpenos que axudan na captación de luz na fotosíntese. As ceras teñen un papel protector debido ao seu carácter hidrófobo.
Metabolismo
[editar | editar a fonte]Os principais lípidos da dieta humana e doutros animais son os triglicéridos de orixe animal e vexetal, esterois, e fosfolípidos de membrana. No metabolismo dos lípidos sintetízanse e degrádanse os lípidos almacenados e prodúcense os lípidos estruturais e funcionais característicos de cada tecido.
Biosíntese
[editar | editar a fonte]Nos animais, cando hai unha inxesta excesiva de carbohidratos na dieta, o exceso de carbohidratos convértese en triglicéridos. Isto implica a biosíntese de ácidos graxos a partir de acetil-CoA e a esterificación dos ácidos graxos para a produción dos triglicéridos, un proceso chamado lipoxénese.[73] Os ácidos graxos son producidos polas ácido graxo sintases, que polimerizan e despois reducen as unidades de acetil-CoA. As cadeas hidrocarbonadas dos ácidos graxos alónganse mediante unhas reaccións cíclicas que engaden o grupo acetilo, redúceno a alcohol, deshidrátano a un grupo alqueno e despois redúceno outra vez a un grupo alcano. Os encimas que interveñen na biosíntese de ácidos graxos divídense en dous grupos, en animais e fungos todas estas reaccións da ácido graxo sintase lévanas a cabo un conxunto unido multifuncional de proteínas,[74] mentres que nos plastos das plantas e nas bacterias son encimas separados os que realizan cada paso da vía biosintética.[75][76] Os ácidos graxos poden ser posteriormente convertidos en triglicéridos que se empaquetan en lipoproteínas e son segregados no fígado.
A síntese de ácidos graxos insaturados require unha reacción de desaturación na cal se introduce un dobre enlace na cadea de acilo graxo. Por exemplo, nos humanos, a desaturación de ácido esteárico pola estearoíl-CoA desaturase-1 produce ácido oleico. O ácido graxo con dous enlaces insaturados ácido linoleico e o de tres enlaces insaturados ácido α-linolénico non poden sintetizarse nos tecidos dos mamíferos, e denomínanse ácidos graxos esenciais e deben obterse da dieta.[77]
A síntese de triglicéridos ten lugar no retículo endoplasmático por vías metabólicas nas cales os grupos acilo dos acilo graxo-CoAs se transfiren aos grupos hidroxilo do glicerol 3-fosfato e do diacilglicerol.[78]
Os terpenos e isoprenoides, incluíndo os carotenoides, están formados pola ensamblaxe e modificación de unidades de isopreno doadas polos precursores reactivos pirofosfato de isopentenilo e pirofosfato de dimetilalilo.[79] Estes precursores poden obterse en diferentes vías metabólicas. Nos animais e arqueas, a vía do mevalonato produce estes compostos a partir de acetil-CoA,[80] e nas plantas e bacterias utilízase a vía alternativa á do mevalonato que usa piruvato e gliceraldehido 3-fosfato como substratos.[79][81] Unha reacción importante que utiliza estes doantes de isopreno activados é a biosíntese de esteroides. Aquí, enlázanse as unidades de isopreno para fabricar escualeno e despois este dóbrase e forma un conxunto de aneis na molécula converténdose en lanosterol.[82] O lanosterol pode despois converterse noutros esteroides como o colesterol e ergosterol.[82][83]
Degradación
[editar | editar a fonte]A beta-oxidación dos ácidos graxos é o proceso metabólico polo cal se degradan os ácidos graxos, e ten lugar na mitocondria e/ou nos peroxisomas para xerar acetil-CoA. Esencialmente, os ácidos graxos oxídanse por un mecanismo que é similar, pero non idéntico, ao inverso da síntese de ácidos graxos, e que consiste en que se eliminan fragmentos de dous carbonos secuencialmente do extremo carboxilo do ácido graxo realizando reaccións de deshidroxenación, hidratación, e oxidación para formar un beta-cetoácido, que se separa por tiólise. O acetil-CoA é despois finalmente convertido en ATP, CO2, e H2O utilizando o ciclo do ácido cítrico ou de Krebs e a cadea de transporte de electróns.
O ciclo de Krebs pode iniciarse con este acetil-CoA producido pola degradación das graxas, que é utilizado para a produción de enerxía cando non hai ou hai pouca glicosa dispoñible.
A enerxía obtida na oxidación completa do ácido graxo palmitato é de 106 ATP.[84] Os ácidos graxos insaturados e os de cadea impar requiren pasos encimáticos adicionais para a súa degradación.
Nutrición e saúde
[editar | editar a fonte]A maioría das graxas que se encontran nos alimentos son triglicéridos, colesterol, e fosfolípidos. Algunhas graxas da nosa dieta son necesarias para facilitar a absorción de vitaminas liposolubles (A, D, E, e K) e carotenoides.[85] Os humanos e outros animais teñen un requirimento de ácidos graxos esenciais que se deben tomar na dieta como o ácido linoleico (un ácido graxo omega-6) e o ácido alfa-linolénico (un ácido graxo omega-3) porque non poden ser sintetizados no organismo a partir de precursores simples.[77] Estes dous ácidos graxos esenciais son de 18 carbonos e poliinsaturados e diferéncianse no número e posición dos seus dobres enlaces. A maioría dos aceites vexetais son ricos en ácido linoleico (aceite de cártamo, aceite de xirasol, e aceite de millo). O ácido alfa linolénico encóntrase nas follas verdes de moitas plantas e en sementes, noces e legumes (especialmente no aceite de liñaza, colza, noces e soia).[86] Os aceites de peixe son especialmente ricos en ácidos graxos omega-3 de cadea máis longa como o ácido eicosapentaenoico (EPA) e ácido docosahexaenoico (DHA).[87] Un gran número de estudos mostraron os beneficios para a saúde asociados co consumo de ácidos graxos omega-3 no desenvolvemento dos nenos, cancro, doenzas cardiovasculares, e varias doenzas mentais, como a depresión, trastorno por déficit de atención con hiperactividade, e demencia.[88][89] Polo contrario, non está ben establecido que o consumo de graxas trans, como as presentes nos aceites vexetais parcialmente hidroxenados, sexan un factor de risco para as doenzas cardiovasculares.[90][91][92]
Uns poucos estudos suxeriron que o consumo de graxas na dieta total está ligado a un incremento do risco de obesidade[93][94] e diabetes.[95][96] Porén, varios estudos moi amplos, incluíndo o Women's Health Initiative Dietary Modification Trial, que estudou a 49.000 mulleres e durou oito anos, non atoparon tal relación.[97][98][99] Ningún deses amplos estudos suxeriu ningunha conexión entre a porcentaxe de calorías procedentes das graxas e o risco de cancro, enfermidades do corazón ou aumento de peso. O Departamento de Nutrición da Escola de Harvard de Saúde Pública mantén unha páxina web que resume as evidencias actuais sobre o impacto das graxas da dieta na saúde, onde se pode ler: "As investigacións detalladas —moitas das cales foron feitas en Harvard — indican que a cantidade total de graxas na dieta non está realmente ligada co peso ou a enfermidade. O que realmente importa é o tipo de graxas e o total de calorías na dieta. As graxas malas, que son as trans e as saturadas, incrementan o risco de ter certas enfermidades. As graxas boas, que son as monounsaturadas e poliinsaturadas, fan xusto o contrario."[100]
Notas
[editar | editar a fonte]- ↑ Maitland, Jr Jones (1998). Organic Chemistry. W W Norton & Co Inc (Np). p. 139. ISBN 0-393-97378-6.
- ↑ Stryer et al., p. 328.
- ↑ 3,0 3,1 3,2 Stryer et al., p. 330.
- ↑ 4,0 4,1 4,2 4,3 Fahy E, Subramaniam S, Murphy R, Nishijima M, Raetz C, Shimizu T, Spener F, Van Meer G, Wakelam M and Dennis E.A (2009). "Update of the LIPID MAPS comprehensive classification system for lipids". Journal of Lipid Research 50 (Suplemento): S9–S14. PMC 2674711. PMID 19098281. doi:10.1194/jlr.R800095-JLR200.
- ↑ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes RW, Cotter D, Dinasarapu AR and Maurya MR (2011). "Bioinformatics and Systems Biology of the Lipidome". Chemical Reviews 111 (10): 6452–6490. PMC 3383319. PMID 21939287. doi:10.1021/cr200295k.
- ↑ Mashaghi S., Jadidi T., Koenderink G., Mashaghi A. (2013). "Lipid Nanotechnology". Int. J. Mol. Sci. 2013 (14): 4242–4282. doi:10.3390/ijms14024242.
- ↑ Michelle A, Hopkins J, McLaughlin CW, Johnson S, Warner MQ, LaHart D, Wright JD (1993). Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall. ISBN 0-13-981176-1. OCLC 32308337.
- ↑ Vance JE, Vance DE (2002). Biochemistry of Lipids, Lipoproteins and Membranes. Amsterdam: Elsevier. ISBN 0-444-51139-3. OCLC 51001207.
- ↑ Brown HA, ed. (2007). Lipodomics and Bioactive Lipids: Mass Spectrometry Based Lipid Analysis, Volume 432 (Methods in Enzymology). Boston: Academic Press. ISBN 0-12-373895-4. OCLC 166624879.
- ↑ "Types of Lipids".
- ↑ Hunt SM, Groff JL, Gropper SAS (1995). West Pub. Co, ed. Advanced Nutrition and Human Metabolism. Belmont, CA. p. 98. ISBN 0-314-04467-1.
- ↑ Devlin, pp. 193–95.
- ↑ Hunter JE (2006). "Dietary trans fatty acids: review of recent human studies and food industry responses". Lipids 41 (11): 967–92. PMID 17263298. doi:10.1007/s11745-006-5049-y.
- ↑ "A Long Lipid, a Long Name: Docosahexaenoic Acid". The Lipid Chronicles. Consultado o 2011-12-31.
- ↑ DHA/EPA Omega-3 Institute (ed.). "DHA FOR OPTIMAL BRAIN AND VISUAL FUNCTIONING". Arquivado dende o orixinal o 10 de marzo de 2016. Consultado o 02 de maio de 2013.
- ↑ Fezza F, De Simone C, Amadio D, Maccarrone M (2008). "Fatty acid amide hydrolase: a gate-keeper of the endocannabinoid system". Subcellular Biochemistry. Subcellular Biochemistry 49: 101–32. ISBN 978-1-4020-8830-8. PMID 18751909. doi:10.1007/978-1-4020-8831-5_4.
- ↑ Coleman RA, Lee DP (2004). "Enzymes of triglyceride synthesis and their regulation". Progress in Lipid Research 43 (2): 134–76. PMID 14654091. doi:10.1016/S0163-7827(03)00051-1.
- ↑ van Holde and Mathews, p. 630–31.
- ↑ 19,0 19,1 Hölzl G, Dörmann P (2007). "Structure and function of glycoglycerolipids in plants and bacteria". Progress in Lipid Research 46 (5): 225–43. PMID 17599463. doi:10.1016/j.plipres.2007.05.001.
- ↑ Honke K, Zhang Y, Cheng X, Kotani N, Taniguchi N (2004). "Biological roles of sulfoglycolipids and pathophysiology of their deficiency" (PDF). Glycoconjugates Journal 21 (1–2): 59–62. PMID 15467400. doi:10.1023/B:GLYC.0000043749.06556.3d. Consultado o 15 de marzo de 2022.
- ↑ "The Structure of a Membrane". The Lipid Chronicles. Consultado o 2011-12-31.
- ↑ Berridge MJ, Irvine RF (1989). "Inositol phosphates and cell signalling" (PDF). Nature Journal 341 (1): 197–205. doi:10.1038/341197a0.
- ↑ Farooqui AA, Horrocks LA, Farooqui T (2000). "Glycerophospholipids in brain: their metabolism, incorporation into membranes, functions, and involvement in neurological disorders". Chemistry and Physics of Lipids 106 (1): 1–29. PMID 10878232. doi:10.1016/S0009-3084(00)00128-6. Consultado o 2009-04-12.
- ↑ Ivanova PT, Milne SB, Byrne MO, Xiang Y, Brown HA (2007). "Glycerophospholipid identification and quantitation by electrospray ionization mass spectrometry". Methods in Enzymology. Methods in Enzymology 432: 21–57. ISBN 9780123738950. PMID 17954212. doi:10.1016/S0076-6879(07)32002-8.
- ↑ van Holde and Mathews, p. 844.
- ↑ Paltauf F (1994). "Ether lipids in biomembranes". Chemistry and Physics of Lipids 74 (2): 101–39. PMID 7859340. doi:10.1016/0009-3084(94)90054-X.
- ↑ Merrill AH, Sandhoff K. (2002). "Sphingolipids: metabolism and cell signaling", in New Comprehensive Biochemistry: Biochemistry of Lipids, Lipoproteins,and Membranes, Vance, D.E. and Vance, J.E., eds. Elsevier Science, NY. Ch. 14.
- ↑ Devlin, pp. 421–22.
- ↑ Hori T, Sugita M (1993). "Sphingolipids in lower animals". Prog. Lipid Res 32 (1): 25–45. PMID 8415797. doi:10.1016/0163-7827(93)90003-F.
- ↑ Wiegandt H (1992). "Insect glycolipids". Biochimica et Biophysica Acta 1123 (2): 117–26. PMID 1739742. doi:10.1016/0005-2760(92)90101-Z.
- ↑ Guan X, Wenk MR (2008). "Biochemistry of inositol lipids". Frontiers in Bioscience 13 (13): 3239–51. PMID 18508430. doi:10.2741/2923.
- ↑ Bach D, Wachtel E (2003). "Phospholipid/cholesterol model membranes: formation of cholesterol crystallites". Biochim Biophys Acta 1610 (2): 187–97. PMID 12648773. doi:10.1016/S0005-2736(03)00017-8.
- ↑ Stryer et al., p. 749.
- ↑ Bouillon R, Verstuyf A, Mathieu C, Van Cromphaut S, Masuyama R, Dehaes P, Carmeliet G (2006). "Vitamin D resistance". Best Practice & Research. Clinical Endocrinology & Metabolism 20 (4): 627–45. PMID 17161336. doi:10.1016/j.beem.2006.09.008.
- ↑ Russell DW (2003). "The enzymes, regulation, and genetics of bile acid synthesis". Annual Review of Biochemistry 72: 137–74. PMID 12543708. doi:10.1146/annurev.biochem.72.121801.161712.
- ↑ Villinski JC, Hayes JM, Brassell SC, Riggert VL, Dunbar RB (2008). "Sedimentary sterols as biogeochemical indicators in the Southern Ocean". Organic Geochemistry 39 (5): 567–88. doi:10.1016/j.orggeochem.2008.01.009.
- ↑ Deacon J (2005). Fungal Biology. Cambridge, MA: Blackwell Publishers. p. 342. ISBN 1-4051-3066-0.
- ↑ Kuzuyama T, Seto H (2003). "Diversity of the biosynthesis of the isoprene units". Natural Product Reports 20 (2): 171–83. PMID 12735695. doi:10.1039/b109860h.
- ↑ Rao AV, Rao LG (2007). "Carotenoids and human health". Pharmacological Research : the Official Journal of the Italian Pharmacological Society 55 (3): 207–16. PMID 17349800. doi:10.1016/j.phrs.2007.01.012.
- ↑ Brunmark A, Cadenas E (1989). "Redox and addition chemistry of quinoid compounds and its biological implications". Free Radical Biology & Medicine 7 (4): 435–77. PMID 2691341. doi:10.1016/0891-5849(89)90126-3.
- ↑ Swiezewska E, Danikiewicz W (2005). "Polyisoprenoids: structure, biosynthesis and function". Progress in Lipid Research 44 (4): 235–58. PMID 16019076. doi:10.1016/j.plipres.2005.05.002.
- ↑ 42,0 42,1 Raetz CR, Garrett TA, Reynolds CM, Shaw WA, Moore JD, Smith DC Jr, Ribeiro AA, Murphy RC,Ulevitch RJ, Fearns C, Reichart D, Glass CK, Benner C, Subramaniam S, Harkewicz R, Bowers-Gentry RC, Buczynski MW, Cooper JA, Deems RA, Dennis EA (2006). "Kdo2-Lipid A of Escherichia coli, a defined endotoxin that activates macrophages via TLR-4". Journal of Lipid Research 47 (5): 1097–111. PMID 16479018. doi:10.1194/jlr.M600027-JLR200.
- ↑ Walsh CT (2004). "Polyketide and nonribosomal peptide antibiotics: modularity and versatility". Science 303 (5665): 1805–10. PMID 15031493. doi:10.1126/science.1094318.
- ↑ Caffrey P, Aparicio JF, Malpartida F, Zotchev SB (2008). "Biosynthetic engineering of polyene macrolides towards generation of improved antifungal and antiparasitic agents". Current Topics in Medicinal Chemistry 8 (8): 639–53. PMID 18473889. doi:10.2174/156802608784221479. Consultado o 2009-04-12.
- ↑ Minto RE, Blacklock BJ (2008). "Biosynthesis and function of polyacetylenes and allied natural products". Progress in Lipid Research 47 (4): 233–306. PMC 2515280. PMID 18387369. doi:10.1016/j.plipres.2008.02.002. Consultado o 2009-04-12.
- ↑ Stryer et al., pp. 329–331
- ↑ Heinz E.(1996). Plant glycolipids: structure, isolation and analysis. in Advances in Lipid Methodology - 3, pp. 211–332 (ed. W.W. Christie, Oily Press, Dundee)
- ↑ Stryer et al., pp. 333–34.
- ↑ van Meer G, Voelker DR, Feigenson GW (2008). "Membrane lipids: where they are and how they behave". Nature Reviews Molecular Cell Biology 9 (2): 112–24. PMC 2642958. PMID 18216768. doi:10.1038/nrm2330.
- ↑ Feigenson GW (2006). "Phase behavior of lipid mixtures". Nature Chemical Biology 2 (11): 560–63. PMC 2685072. PMID 17051225. doi:10.1038/nchembio1106-560.
- ↑ Wiggins PM (1990). "Role of water in some biological processes". Microbiological Reviews 54 (4): 432–49. PMC 372788. PMID 2087221.
- ↑ Raschke TM, Levitt M (2005). "Nonpolar solutes enhance water structure within hydration shells while reducing interactions between them". Proceedings of the National Academy of Sciences U.S.A 102 (19): 6777–82. PMC 1100774. PMID 15867152. doi:10.1073/pnas.0500225102.
- ↑ Brasaemle DL (2007). "Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis". J. Lipid Res 48 (12): 2547–59. PMID 17878492. doi:10.1194/jlr.R700014-JLR200. Arquivado dende o orixinal o 06 de xullo de 2010. Consultado o 02 de maio de 2013.
- ↑ Stryer et al., p. 619.
- ↑ "Copia arquivada". Arquivado dende o orixinal o 23 de outubro de 2011. Consultado o 02 de maio de 2013.
- ↑ Wang X (2004). "Lipid signaling". Current Opinions in Plant Biology 7 (3): 329–36. PMID 15134755. doi:10.1016/j.pbi.2004.03.012.
- ↑ Dinasarapu AR,Saunders B,Ozerlat I,Azam K, and Subramaniam S (2011). "Signaling gateway molecule pages—a data model perspective". Bioinformatics 27 (12): 1736–1738. PMC 3106186. PMID 21505029. doi:10.1093/bioinformatics/btr190.
- ↑ Eyster KM (2007). "The membrane and lipids as integral participants in signal transduction". Advances in Physiology Education 31 (1): 5–16. PMID 17327576. doi:10.1152/advan.00088.2006.
- ↑ Hinkovska-Galcheva V, VanWay SM, Shanley TP, Kunkel RG (2008). "The role of sphingosine-1-phosphate and ceramide-1-phosphate in calcium homeostasis". Current Opinion in Investigational Drugs 9 (11): 1192–205. PMID 18951299.
- ↑ Saddoughi SA, Song P, Ogretmen B (2008). "Roles of bioactive sphingolipids in cancer biology and therapeutics". Subcellular Biochemistry. Subcellular Biochemistry 49: 413–40. ISBN 978-1-4020-8830-8. PMC 2636716. PMID 18751921. doi:10.1007/978-1-4020-8831-5_16.
- ↑ Klein C, Malviya AN (2008). "Mechanism of nuclear calcium signaling by inositol 1,4,5-trisphosphate produced in the nucleus, nuclear located protein kinase C and cyclic AMP-dependent protein kinase". Frontiers in Bioscience 13 (13): 1206–26. PMID 17981624. doi:10.2741/2756.
- ↑ Boyce JA (2008). "Eicosanoids in asthma, allergic inflammation, and host defense". Current Molecular Medicine 8 (5): 335–49. PMID 18691060. doi:10.2174/156652408785160989.
- ↑ Bełtowski J (2008). "Liver X receptors (LXR) as therapeutic targets in dyslipidemia". Cardiovascular Therapy 26 (4): 297–316. PMID 19035881. doi:10.1111/j.1755-5922.2008.00062.x.
- ↑ Yi Wu, Nitu Tibrewal and Raymond B. Birge. Phosphatidylserine recognition by phagocytes: a view to a kill. Trends in Cell Biology, Volume 16, Issue 4, 189-197, 1 April 2006. doi 10.1016/j.tcb.2006.02.003. [1]
- ↑ Callahan MK, Williamson P, Schlegel RA. Surface expression of phosphatidylserine on macrophages is required for phagocytosis of apoptotic thymocytes. Cell Death Differ. 2000 Jul;7(7):645-53. PMID 10889509. [2]
- ↑ Indiveri C, Tonazzi A, Palmieri F (1991). "Characterization of the unidirectional transport of carnitine catalyzed by the reconstituted carnitine carrier from rat liver mitochondria". Biochim. Biophys. Acta 1069 (1): 110–6. PMID 1932043. doi:10.1016/0005-2736(91)90110-T.
- ↑ Parodi AJ, Leloir LF (1979). "The role of lipid intermediates in the glycosylation of proteins in the eucaryotic cell". Biochim. Biophys. Acta 559 (1): 1–37. PMID 375981. doi:10.1016/0304-4157(79)90006-6.
- ↑ Helenius A, Aebi M (2001). "Intracellular functions of N-linked glycans". Science 291 (5512): 2364–69. PMID 11269317. doi:10.1126/science.291.5512.2364.
- ↑ M. Nowicki and M. Frentzen (2005). "Cardiolipin synthase of Arabidopsis thaliana". FEBS Letters 579 (10): 2161–2165. PMID 15811335. doi:10.1016/j.febslet.2005.03.007.
- ↑ M. Nowicki (2006). "Characterization of the Cardiolipin Synthase from Arabidopsis thaliana". Ph.D. thesis, RWTH-Aachen University. Arquivado dende o orixinal o 05 de outubro de 2011. Consultado o 02 de maio de 2013.
- ↑ Gohil VM, Greenberg ML (2009). "Mitochondrial membrane biogenesis: phospholipids and proteins go hand in hand". Journal of Cell Biology 184 (4): 469–72. PMC 2654137. PMID 19237595. doi:10.1083/jcb.200901127.
- ↑ Hoch FL (1992). "Cardiolipins and biomembrane function". Biochimica et Biophysica Acta 1113 (1): 71–133. PMID 10206472. doi:10.1016/0304-4157(92)90035-9.
- ↑ Stryer et al., p. 634.
- ↑ Chirala S, Wakil S (2004). "Structure and function of animal fatty acid synthase". Lipids 39 (11): 1045–53. PMID 15726818. doi:10.1007/s11745-004-1329-9.
- ↑ White S, Zheng J, Zhang Y (2005). "The structural biology of type II fatty acid biosynthesis". Annual Review of Biochemistry 74: 791–831. PMID 15952903. doi:10.1146/annurev.biochem.74.082803.133524.
- ↑ Ohlrogge J, Jaworski J (1997). "Regulation of fatty acid synthesis". Annual Review of Plant Physiology and Plant Molecular Biology 48: 109–136. PMID 15012259. doi:10.1146/annurev.arplant.48.1.109.
- ↑ 77,0 77,1 Stryer et al., p. 643.
- ↑ Stryer et al., pp. 733–39.
- ↑ 79,0 79,1 Kuzuyama T, Seto H (2003). "Diversity of the biosynthesis of the isoprene units". Natural Product Reports 20 (2): 171–83. PMID 12735695. doi:10.1039/b109860h.
- ↑ Grochowski L, Xu H, White R (2006). "Methanocaldococcus jannaschii uses a modified mevalonate pathway for biosynthesis of isopentenyl diphosphate". Journal of Bacteriology 188 (9): 3192–98. PMC 1447442. PMID 16621811. doi:10.1128/JB.188.9.3192-3198.2006.
- ↑ Lichtenthaler H (1999). "The 1-Dideoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants". Annual Review of Plant Physiology and Plant Molecular Biology 50: 47–65. PMID 15012203. doi:10.1146/annurev.arplant.50.1.47.
- ↑ 82,0 82,1 Schroepfer G (1981). "Sterol biosynthesis". Annual Review of Biochemistry 50: 585–621. PMID 7023367. doi:10.1146/annurev.bi.50.070181.003101.
- ↑ Lees N, Skaggs B, Kirsch D, Bard M (1995). "Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review". Lipids 30 (3): 221–26. PMID 7791529. doi:10.1007/BF02537824.
- ↑ Stryer et al., pp. 625–26.
- ↑ Bhagavan, p. 903.
- ↑ Russo GL (2009). "Dietary n-6 and n-3 polyunsaturated fatty acids: from biochemistry to clinical implications in cardiovascular prevention". Biochemical Pharmacology 77 (6): 937–46. PMID 19022225. doi:10.1016/j.bcp.2008.10.020.
- ↑ Bhagavan, p. 388.
- ↑ Riediger ND, Othman RA, Suh M, Moghadasian MH (2009). "A systemic review of the roles of n-3 fatty acids in health and disease". Journal of the American Dietetic Association 109 (4): 668–79. PMID 19328262. doi:10.1016/j.jada.2008.12.022.
- ↑ Galli C, Risé P (2009). "Fish consumption, omega 3 fatty acids and cardiovascular disease. The science and the clinical trials". Nutrition and Health (Berkhamsted, Hertfordshire) 20 (1): 11–20. PMID 19326716. doi:10.1177/026010600902000102.
- ↑ Micha R, Mozaffarian D (2008). "Trans fatty acids: effects on cardiometabolic health and implications for policy". Prostaglandins, Leukotrienes, and Essential Fatty Acids 79 (3–5): 147–52. PMC 2639783. PMID 18996687. doi:10.1016/j.plefa.2008.09.008.
- ↑ Dalainas I, Ioannou HP (2008). "The role of trans fatty acids in atherosclerosis, cardiovascular disease and infant development". International Angiology: a Journal of the International Union of Angiology 27 (2): 146–56. PMID 18427401.
- ↑ Mozaffarian D, Willett WC (2007). "Trans fatty acids and cardiovascular risk: a unique cardiometabolic imprint?". Current Atherosclerosis Reports 9 (6): 486–93. PMID 18377789. doi:10.1007/s11883-007-0065-9.
- ↑ Astrup A, Dyerberg J, Selleck M, Stender S (2008). "Nutrition transition and its relationship to the development of obesity and related chronic diseases". Obesity Review. 9 Suppl 1: 48–52. PMID 18307699. doi:10.1111/j.1467-789X.2007.00438.x.
- ↑ Astrup A (2005). "The role of dietary fat in obesity". Seminars in Vascular Medicine 5 (1): 40–47. PMID 15968579. doi:10.1055/s-2005-871740.
- ↑ Ma Y.; Olendzki, Barbara C.; Hafner, Andrea R.; Chiriboga, David E.; Culver, Annie L.; Andersen, Victoria A.; Merriam, Philip A.; Pagoto, Sherry L.; et al. (2006). "Low-carbohydrate and high-fat intake among adult patients with poorly controlled type 2 diabetes mellitus". Nutrition 22 (11-12): 1129–1136. PMC 2039705. PMID 17027229. doi:10.1016/j.nut.2006.08.006.
- ↑ Astrup A (2008). "Dietary management of obesity". JPEN Journal of Parenteral and Enteral Nutrition 32 (5): 575–77. PMID 18753397. doi:10.1177/0148607108321707.
- ↑ Beresford SA, Johnson KC, Ritenbaugh C; et al. (2006). "Low-fat dietary pattern and risk of colorectal cancer: the Women's Health Initiative Randomized Controlled Dietary Modification Trial". JAMA: the Journal of the American Medical Association 295 (6): 643–54. PMID 16467233. doi:10.1001/jama.295.6.643.
- ↑ Howard BV, Manson JE, Stefanick ML; et al. (2006). "Low-fat dietary pattern and weight change over 7 years: the Women's Health Initiative Dietary Modification Trial". JAMA: the Journal of the American Medical Association 295 (1): 39–49. PMID 16391215. doi:10.1001/jama.295.1.39.
- ↑ Howard BV, Van Horn L, Hsia J; et al. (2006). "Low-fat dietary pattern and risk of cardiovascular disease: the Women's Health Initiative Randomized Controlled Dietary Modification Trial". JAMA : the Journal of the American Medical Association 295 (6): 655–66. PMID 16467234. doi:10.1001/jama.295.6.655.
- ↑ "Fats and Cholesterol: Out with the Bad, In with the Good — What Should You Eat? - The Nutrition Source — Harvard School of Public Health". Arquivado dende o orixinal o 27 de abril de 2011. Consultado o 4 maio 2013.
Véxase tamén
[editar | editar a fonte]| Commons ten máis contidos multimedia sobre: Lípido |
Bibliografía
[editar | editar a fonte]- Bhagavan NV (2002). Medical Biochemistry. San Diego: Harcourt/Academic Press. ISBN 0-12-095440-0.
- Devlin TM (1997). Textbook of Biochemistry: With Clinical Correlations (4th ed.). Chichester: John Wiley & Sons. ISBN 0-471-17053-4.
- Stryer L, Berg JM, Tymoczko JL (2007). Biochemistry (6th ed.). San Francisco: W.H. Freeman. ISBN 0-7167-8724-5.
- Van Holde KE, Mathews CK (1996). Biochemistry (2nd ed.). Menlo Park, Calif: Benjamin/Cummings Pub. Co. ISBN 0-8053-3931-0.
Outros artigos
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]Introdutorias
- List de webs relacionadas con lípidos
- Nature Lipidomics Gateway - Round-up and summaries of recent lipid research
- Lipid Library - Referencias xerais da química e bioquímica de lípidos
- Cyberlipid.org - Recursos sobre a historia dos lípidos.
- Molecular Computer Simulations - Modelos de membranas lipídicas
- Tráfico de lípidos, membranas e vesículas - Libraría virtual de bioquímica e bioloxía celular
Nomenclatura
Bases de datos
- LIPID MAPS - Xenes e proteínas asociadas a lípidos.
- LipidBank - Base de datos xaponesa de lípidos e propiedades relacionadas, datos espectrais e referencias.
- LIPIDAT Arquivado 25 de febreiro de 2020 en Wayback Machine. - Base de datos principalmente de fosfolípidos e datos termodinámicos asociados.