Pteridophyta
- Para una introducción a estas plantas véase la sección de plantas terrestres en «Planta»
| Helechos y afines | ||
|---|---|---|
 Helechos y afines | ||
 Diagrama esquemático del ciclo de vida de las plantas vasculares sin semilla (Pteridophyta).
Referencias: | ||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| Reino: | Plantae | |
| Subreino: | Tracheobionta | |
| Superdivisión: | Pteridophyta (P) | |
| Divisiones | ||
Según Christenhusz et al. 2011,[1][2][3] basada en Smith et al. 2006,[4] 2008;[5] que también provee una secuencia lineal de las licofitas y monilofitas. | ||
Las pteridofitas, pteridófitas (nombre científico Pteridophyta), criptógamas vasculares o, generalmente, helechos y afines son un grupo de plantas terrestres que se caracterizan por ser traqueofitas con alternancia de generaciones bien manifiesta, donde el esporófito es un cormo primitivo que posee vástago con tallo y generalmente también hojas (microfilos o megafilos) y raíces siempre adventicias, con xilema primitivo compuesto por traqueidas, y floema primitivo compuesto por células cribosas, y en las que el xilema y el floema forman haces vasculares ubicados en un cilindro central rodeado de la corteza primaria o endodermis, rodeada de epidermis con cutícula y estomas, que se mantiene a lo largo de toda la vida del esporófito; sin crecimiento secundario; con esporas como unidad de dispersión que persiste por siglos, formadas en esporangios o leptosporangios; donde el gametófito es un talo (cuerpo sin organización), en el que se forman los arquegonios (órgano sexual pluricelular que originará la gameta femenina inmóvil) y los anteridios (órgano pluricelular donde se formarán las gametas masculinas móviles flageladas), y donde la fecundación es dependiente del agua; con un embrión que se desarrolla sobre el gametófito directamente después de la formación del cigoto y que no entra en latencia, que puede ser bipolar y con suspensor si proviene de un esporangio (posee el meristema apical y el radical, que después no se desarrolla) o unipolar (si proviene de un leptosporangio, solo posee el meristema apical); el embrión crece sobre el gametófito sin ser liberado, y dará el vástago con raíces adventicias.
En general el esporófito es perenne, aunque la parte visible puede desaparecer totalmente en la época desfavorable. El gametófito en cambio en general es efímero, y debe estar ligado al agua para su subsistencia y para que la reproducción sea exitosa. La presión de selección actúa principalmente sobre el esporófito, estando el gametófito más conservado evolutivamente. La reproducción vegetativa puede darse por propágulos del esporófito, es especialmente exitosa en pteridofitos colonizadores.
Las esporas son la unidad de dispersión y las responsables de colonizar nuevos hábitats (en las gimnospermas es la semilla, en las angiospermas es el fruto), y también constituyen las unidades de resistencia en las épocas desfavorables (en las espermatofitas es la semilla con el embrión latente).
Las pteridofitas son las más antiguas de las traqueofitas actuales, ya existían en el Devónico y tuvieron su apogeo a fines del Mesozoico. Las pteridofitas actuales comprenden cerca de 12 000 especies concentradas en áreas tropicales con diversidad de ambientes, y áreas montañosas húmedas. Su importancia es sistemática. De poca importancia económica, se las cultiva principalmente como ornamentales.
Los análisis moleculares de ADN muestran que las pteridofitas son un grupo parafilético compuesto por dos grupos monofiléticos distintos: Lycophyta, que contiene a las pteridofitas más antiguas con microfilos, y Monilophyta, que reúne a las pteridofitas con megafilos y a las que perdieron sus megafilos luego de haberlos adquirido. Los dos taxones son dos eslabones evolutivos intermedios entre Bryophyta y Spermatophyta (Pryer et al. 2001, Judd et al. 2002, Pryer et al. 2004a).
Diversidad
[editar]Esta es una lista de los representantes más definidos de cada grupo de pteridofitos, creada con el fin de mostrar la diversidad necesaria para comprender los avances en taxonomía y filogenia de pteridófitos. Las descripciones de cada grupo son deliberadamente incompletas. Para una lista completa de taxones, o una explicación más detallada de los mismos, siga el árbol taxonómico en la ficha presentada al principio de esta página, o ingrese al artículo principal.
Lycopodium y afines
[editar]Los esporófitos de las licopodiáceas que viven en la actualidad son de porte pequeño y con licofilos libres (no soldados entre sí). La parte del tallo con licofilos esporangiados forma el "estróbilo".
Carácter de importancia taxonómica:. Los microfilos son "licofilos". Los esporangios se ubican en la cara adaxial de los licofilos. Poseen esporangios de tipo eusporangiado, de dehiscencia transversal, típicamente reniforme (con forma de riñón).
-
Figura 4.2. Lycopodiaceae. Nótense los licofilos estériles en la parte superior, y los estróbilos de licofilos fértiles en la parte inferior.
-
Figura 4.3. Estróbilo de Selaginella preservado en líquido. Nótese que los esporangios están ubicados en la cara adaxial de los esporofilos.



Equisetum
[editar]Equisetum o "cola de caballo" es un esporófito con rizoma subterráneo del que se originan tallos aéreos, con hojas pequeñas, escamosas, dispuestas en el tallo en verticilos, en cada verticilo las hojas están soldadas entre sí. Al principio son fotosintéticas pero luego se secan. La principal función de fotosíntesis es realizada por el tallo verde, con cloroplastos. Los esporangios se ubican en las hojas fértiles o "esporofilos", no fotosintéticas y peltadas (unidas al tallo por un pie), ubicadas en el sector apical del tallo llamado "estróbilo". El tallo posee crestas y valles muy característicos, con sílice.
Carácter de importancia taxonómica: Los esporangios son de tipo eusporangiado. El gametófito es autótrofo fotosintético. La característica morfología del tallo hace que sea fácil determinar sus parientes fósiles, entre los que se encuentran equisetos arborescentes de porte impresionante.

Psilotum y afines
[editar]Psilotum no tiene raíces, solo posee un tallo subterráneo. Sus hojas están tan reducidas que durante mucho tiempo se consideró que no poseía hojas. Los esporangios están agrupados en sinangios que se ubican directamente sobre el tallo, y parecen nacer del lado adaxial de las pequeñas hojas bifurcadas. Es curiosa la ramificación del tallo, del tipo dicotómico (el tallo se subdivide en dos ramas iguales, que a su vez pueden dividirse en dos ramas iguales, y así). Si bien la reducción de sus caracteres (como raíces y hojas) fue adquirida de forma secundaria, esta característica y la ramificación dicotómica durante mucho tiempo lo hizo parecer emparentado a antiguos fósiles de plantas vasculares como Rhynia.
Junto con otro género forma la familia de las psilotáceas.
Carácter de importancia taxonómica: Los esporangios son de tipo eusporangiado. A diferencia de la mayoría de las pteridofitas el gametofito es axial, subterráneo y micorrícico.
Ophioglossum, Botrychium y afines
[editar]Poseen esporófitos con eufilos no reducidos, son los únicos pteridofitos con megafilos con prefoliación plegada en lugar de circinada. Poseen una sola hoja (fronde) por cada estación de crecimiento. La fronde consta de un sector basal, fértil, y uno apical, que es la lámina fotosintética (en Ophioglossum el sector apical y el basal son enteros, y en Botrychium los dos son divididos). Al sector fértil se lo conoce como "esporóforo", y es fácil apreciar los esporangios sobre él. Se los encuentra en márgenes de cuerpo de agua o terrenos de agua permanente.
Carácter de importancia taxonómica: Poseen esporangios de tipo eusporangiado, agrupados a lo largo del esporoforo. Poseen raíces sin pelos radicales, micorrícicas (asociadas con hongos). A diferencia de la mayoría de las pteridofitas, poseen gametofito axial subterráneo heterótrofo micotrófico, no fotosintético.
-
Figura 4.5. Ophioglossum vulgatum. Nótese la fronde con una pina basal fértil y una pina apical estéril.
-
Figura 4.6. Botrychium lunaria. En esta planta joven puede observarse la prefoliación plegada característica de las ophioglossaceae.


Marattia y afines
[editar]Todos los helechos a partir de aquí son con megafilos (frondes) de prefoliación circinada.
Las marattiáceas más características son helechos de frondes de gran porte, pinadas, son plantas adaptadas para crecer en el suelo sombreado de las selvas tropicales húmedas. Tienen estípulas (evaginaciones de la epidermis alrededor del pecíolo de las hojas), por lo que son fácilmente reconocibles. Angiopteris es conocida en los jardines botánicos que la consideran atractiva debido a sus frondes de gran porte.
Carácter de importancia taxonómica: El esporófito posee un rizoma con una estela característica, única, llamada "polistela" (o también "dictiostela polistélica"). Los esporangios son de tipo eusporangiado.
-
Figura 4.7. Angiopteris evecta. Trofofilo.
-
Figura 4.8. Angiopteris evecta. Fronde joven, prefoliación circinada.


Osmunda y afines
[editar]Todos los helechos a partir de aquí, además de tener megafilos de prefoliación circinada, tienen esporangios de tipo leptosporangiado.
Las osmundáceas son fácilmente reconocibles por sus trofoesporofilos sectoriales: las frondes tienen algunas pinas fértiles (más pequeñas y no fotosintéticas) y algunas pinas estériles fotosintéticas. Las pinas fértiles se encuentran en la parte apical o en la parte media de la fronde. También tienen estípulas (evaginaciones del tallo alrededor del pecíolo de las hojas), pero no se pueden confundir con las marattiáceas debido a sus característicos trofoesporofilos sectoriales (las marattiáceas tienen los esporangios en la cara abaxial de la fronde, las pinas con esporangios no son reducidas).
Carácter de importancia taxonómica: A diferencia de todas las pteridofitas vistas hasta ahora, los esporangios son de tipo leptosporangiado. Los de las osmundáceas son leptosporangios únicos, no típicos: con anillo de dehiscencia lateral (parche de la pared del esporangio con células de pared engrosada).
-
Figura 4.9. Osmunda regalis. Trofoesporofilos sectoriales.
-
Figura 4.10. Osmunda japonica. Fronde joven, prefoliación circinada.



Hymenophyllum y afines
[editar]Estos helechos son fácilmente reconocibles debido a que sus frondes poseen una lámina monoestratificada (de una sola capa de células de espesor) que les da un aspecto semitransparente. Son epífitos (usan otras plantas más altas como sustrato, sin parasitarlas) o saxícolas (usan las piedras como sustrato), se los encuentra en climas desde tropicales a templado-fríos, pero siempre en bosques con precipitaciones permanentes o selvas nubladas. Absorben agua no solo por las raíces sino por toda la superficie de su cuerpo.
Carácter de importancia taxonómica: Todos los helechos que veremos a partir de aquí tienen anillo que rodea al leptosporangio, lo cual se considera un carácter más avanzado que el leptosporangio con anillo lateral de las osmundáceas. El anillo de las himenofiláceas es transversal-oblicuo. Este tipo de anillo se considera más primitivo que los anillos verticales que veremos después.

Gleichenia y afines
[editar]Las gleicheniáceas son muy fáciles de distinguir debido a sus características frondes divididas pseudodicotómicamente ("falsamente divididas en dos"), lo que es logrado cuando las yemas apicales luego de dar las dos ramas laterales entran en dormición. Son terrestres de hojas coriáceas ("de aspecto de cuero"), a diferencia de muchos helechos no se encuentran en climas tropicales, solo en climas templados a templado-fríos.
Carácter de importancia taxonómica: Poseen leptosporangios con anillo entero (no interrumpido en ninguna parte de su recorrido) oblicuo.
Schizaea, Lygodium, Anemia y afines
[editar]Esta familia es muy antigua y con un largo registro fósil. Sus 3 géneros más sobresalientes (Schizaea, Anemia y Lygodium) difieren mucho entre sí en la morfología del esporófito. Anemia es fácilmente reconocida porque las pinas fértiles son las basales y son muy reducidas, teniendo el aspecto de racimos dorados (por eso en Sudamérica subtropical se la conoce como "doradilla"). Schizaea tiene la particularidad de tener todas sus frondes reducidas ("filiformes"). Lygodium es conocido por ser un helecho trepador, muy apreciado en los jardines de clima cálido de todo el mundo. Lygodium japonicum es un helecho trepador cultivado como ornamental, también considerado invasor en Estados Unidos.
Carácter de importancia taxonómica: El anillo de las schizáceas es entero, de posición transversal subapical, considerado un carácter primitivo dentro de los leptosporangios con anillo.
-
Figura 4.12. Lygodium japonicum. Helecho trepador japonés.
-
Figura 4.13. Esporófito de Anemia mexicana. Este género es conocido en la zona subtropical de Sudamérica como "doradilla". Nótense los trofoesporofilos sectoriales, las pinas basales fértiles reducidas, formando un "racimo" que se eleva por encima de las pinas apicales fotosintéticas.


Marsilea, Salvinia, Azolla y afines
[editar]Prácticamente todos los helechos acuáticos (salvo el género Ceratopteris que es un "helecho típico") pertenecen a este grupo. Marsilea es fácilmente confundida con el "trébol de 4 hojas". Salvinia y Azolla son helechos flotantes, Salvinia es un conocido invasor de diques y aguas estancadas, y Azolla es cultivada en los campos de arroz por su poder fertilizante, adquirido en su relación simbiótica con una bacteria fijadora de nitrógeno. Su característica más sobresaliente como grupo es que poseen un ciclo de vida heterospórico.
Carácter de importancia taxonómica: Poseen leptosporangios sin anillo o con anillo relictual (débilmente marcado, no funcional), por lo que fue perdido en forma secundaria (después de haberse adquirido).
-
Figura 4.15. Marsilea. Pinas apicales fotosintéticas.
-
Figura 4.16. Salvinia natans. Hábito.
-
Figura 4.17. Azolla caroliniana. Hábito.



Cyathea, Dicksonia y afines
[editar]Fácilmente reconocibles debido a su característico tallo formando un estípite que puede llegar a tener varios metros de altura, lo que le da al esporófito un aspecto arbóreo. Casi todos los helechos arborescentes pertenecen a este grupo. Normalmente se los encuentra divididos en varias familias, las más conspicuas son Cyatheaceae ("helechos arborescentes con escamas") y Dicksoniaceae ("helechos arborescentes con pelos"), la diferencia principal es que en las cyatheáceas hay haces vasculares en la médula del tallo (se dice que "la médula está revitalizada").
Carácter de importancia taxonómica: Poseen leptosporangios con anillo vertical completo (no interrumpido en ninguna parte de su recorrido). Según algunos autores (Tyron) debería llamarse anillo oblicuo.
-
Figura 4.18. Dicksonia antarctica. Tallo formando un estípite.
-
Figura 4.19. Cyatheaceae. Hábito. Tallo formando un estípite de porte alto. Nótense las frondes de prefoliación circinada.


Helechos típicos
[editar]Comprende al 80 % de las especies de helechos. Normalmente se los encuentra divididos en una cantidad considerable de familias, entre las que se encuentran Pteridaceae, Aspleniaceae, Polypodiaceae, entre otras. Desde el 2004 (Pryer et al. 2004a), se sabe que son monofiléticos.
Carácter de importancia taxonómica: Todas las especies tienen leptosporangio con anillo vertical incompleto, con estomio. Al deshidratarse el esporangio, el anillo se retrae y el estomio cede liberando las esporas de forma brusca, eso asegura una mejor dispersión de la especie con respecto a aquellas que tienen anillos sin estomio. Por ello se considera que este tipo de leptosporangio es el más avanzado.
-
Figura 4.20. Pteris
-
Figura 4.21. Asplenium
-
Figura 4.22. Polypodium



Morfología: El esporófito de las pteridofitas
[editar]El esporófito es el más importante para la determinación de la planta, al ser más conspicuo y menos conservado evolutivamente. El esporófito es un cormo (cuerpo multicelular organizado en tejidos y órganos, con vástago fotosintético, raíz para absorción de agua y sales, y un sistema de haces vasculares que los vincula), característica que comparte con el resto de las plantas vasculares.[cita requerida]
El vástago del esporófito consta de un tallo y generalmente también de hojas. Las hojas que se originan en el tallo pueden estar o no vascularizadas, en las licofitas no están vascularizadas o están vascularizadas pero no deja trazo foliar en el cilindro vascular, debido a eso son hojas que no pueden crecer mucho y son llamadas licofilos, son características de Lycophyta, la división más primitiva de las pteridofitas. Las modernas hojas vascularizadas, los eufilos, son característicos de la división más moderna Monilophyta, coincidiendo su aparición con la de una sifonostela en el cilindro del tallo, son más desarrollados, son nervados, y al alejarse del tallo dejan una traza foliar y una laguna foliar en el cilindro vascular, que se observan en el corte transversal del tallo (estela). Estas definiciones de hoja (licofilo y eufilo) son en base al origen evolutivo del órgano, por lo que no en todos los casos se va a corresponder su nombre con esta descripción de su morfología. Antiguamente se definía a las hojas según su tamaño y vascularización, y en general coincidían los licofilos con lo que se llamaba "microfilos", y los eufilos con lo que se llamaba "megafilos".[4] Solo hay dos clados de monilofitas que tienen sus eufilos reducidos por haberlos perdido en forma secundaria: Equisetum, entre los que se encuentra la "cola de caballo", y Psilotaceae.[cita requerida]
La raíz embrionaria o radícula típica del cormo puede estar presente en el embrión (en helechos eusporangiados) o no (en helechos leptosporangiados), pero aunque esté presente no se desarrolla. La función de absorción la cumplen las raíces adventicias, que se originan en el vástago.[cita requerida]
El sistema vascular de las pteridofitas es el más primitivo de las traqueofitas actuales: no tiene crecimiento secundario (con la excepción de la licofita Isoetes, que adquirió el crecimiento secundario de forma independiente a las espermatofitas).[cita requerida]
La corteza de las pteridofitas, de origen primario, está compuesta por la endodermis, que es un tejido (primario) característico de estos dos clados (y ausente en la mayoría de las espermatofitas debido a que se pierde durante el crecimiento secundario), y por la epidermis que muchas veces presenta un indumento de tricomas (que pueden ser pelos o escamas) o de pecíolos de hojas muertas, muy útil para la determinación de las familias. Protegiendo a la epidermis se encuentra la cutícula que impermeabiliza las paredes celulares de la epidermis y las protege de la sequedad del ambiente en la vida terrestre, pero también impide el intercambio gaseoso con el ambiente. En la epidermis también se encuentran los estomas (del griego "boca"), poros formados por células que tienen la capacidad de abrirlos o cerrarlos según las condiciones ambientales, que permiten el intercambio gaseoso necesario para la fotosíntesis y la respiración.[cita requerida]
Hacia el interior de la corteza se encuentra el cilindro central, donde se aprecian los haces vasculares, comprendidos por el xilema, que en las pteridofitas es de tipo primitivo, formado por parénquima y traqueidas (vasos cerrados por sus extremos), y el floema, también de tipo primitivo y poco diferenciado, formado por células con numerosas áreas cribosas en sus paredes laterales.[cita requerida]
En el corte transversal del tallo y de la raíz (comúnmente denominado estela) se observa que los haces vasculares de xilema y floema primarios se disponen en el cilindro central según patrones que, a diferencia de las otras traqueofitas, poseen una gran diversidad de tipos y se vuelven útiles para la determinación de las familias. Al ser cormos sin crecimiento secundario, este patrón se mantiene a lo largo de la vida del esporófito adulto.[cita requerida]
En general la estela difiere en un corte transversal del tallo y uno de la raíz; la estela del tallo es la más diversificada y de valor sistemático. Esta diversidad de tipos estelares del tallo se puede agrupar en dos grandes grupos: los derivados de la protostela (los haces se disponen en todo el radio del tallo, son las más primitivas) y los derivados de la sifonostela (tallos con médula —región central sin haces compuesta por diferentes tejidos según la familia—). El tipo de estela más común es derivado de la sifonostela y se llama dictiostela.[cita requerida]
Las raíces adventicias, de origen caulinar, poseen una actinostela (estela con forma de estrella) que puede tener diferente cantidad de radios xilemáticos, que puede ser desde dos (diarcas, forman una estrella de dos puntas) hasta muchos (poliarcas, que forman una estrella de muchas puntas), visibles en un corte transversal. Solo como excepción, hay especies que carecen de raíz (algunas epífitas y algunas acuáticas).[cita requerida]
La reproducción en las pteridofitas
[editar]
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundación
En el tallo o en las hojas del esporófito aparecen los meristemoides que son células con capacidad de dividirse que originarán los esporangios ("estructuras formadoras de esporas"). La ubicación de los esporangios en el vástago es de importancia sistemática. En las licofitas (Lycopodium, Selaginella), los esporangios se ubican sobre los licofilos, en su cara adaxial. En las monilofitas, los esporangios se ubican o en la cara abaxial o en el margen de los eufilos. Las hojas con esporangios se llaman "esporofilos".
Dentro de la pared del esporangio ocurre la meiosis formándose las esporas haploides, primer exponente de la "generación gametofítica". Las esporas se nutren de los tejidos nutricios internos a la pared del esporangio, formando una cavidad en el esporangio a medida que el tejido nutricio es agotado y las esporas maduran. Los esporangios pueden ser solo de 2 tipos: eusporangios (originados de varias células meristemoides, forman una pared del esporangio con varias capas de células y con estomas, más de 2000 esporas por esporangio), o leptosporangios (que dieron origen al grupo monofilético de "helechos leptosporangiados", cada esporangio originado de solo una célula meristemoide, que forma un pie de 1 o 2 células de espesor que lo une al esporófito y una cápsula dentro de la cual ocurre la meiosis y formación de esporas, con una pared del esporangio con una sola capa de células y solo 32 o 64 esporas funcionales por esporangio). Los helechos leptosporangiados son la mayoría de las especies de helechos que hoy viven.
Los esporangios pueden estar dispersos o agrupados en "soros", los soros muchas veces están protegidos por una evaginación de la lámina llamada indusio que puede tener diferentes formas (peltado, cupuliforme, etc), tanto la ubicación y forma de los soros como la forma del indusio son útiles en la determinación de las familias. A veces los esporangios están agrupados y además unidos por tejido parenquimático, formando una unidad a simple vista, entonces son llamados "sinangios", que se ponen en evidencia al ver cortes a la lupa, en los que se aprecian diferentes cavidades, cada una de ellas correspondientes a un esporangio.
Cuando la pared del esporangio se rompe las esporas son liberadas. Tanto el tipo de esporangio (eusporangio, leptosporangio), como la forma de rompimiento de la pared del esporangio o "mecanismo de dehiscencia" (por anillo lateral, por anillo completo, por anillo incompleto con estomio), como la forma de la espora (trilete, monolete; con perisporio o sin perisporio) y los "adornos" que hay en la pared de la espora, son utilizados para la determinación de las familias y especies. Normalmente se necesita un microscopio para ver esas estructuras a la resolución que se necesita para determinarlas.

Las esporas son dispersadas por el viento y donde caen se desarrolla el gametofito, el gametofito debe estar ligado al suelo para que ocurra la fecundación que depende del agua. El gametofito de las pteridofitas es llamado "protalo", normalmente es un talo (cuerpo sin estructurarse en tejidos y órganos) pequeño (menos de 1 cm de diámetro) que absorbe agua por todo el cuerpo pero especialmente por rizoides (evaginaciones alargadas del talo, no son raíces propiamente dichas por no poseer su estructura y sus tejidos). El gametofito de las pteridofitas es de vida libre (no dependiente del esporófito para su nutrición y crecimiento), generalmente es autótrofo, aunque podemos encontrar familias con gametofito saprofito micorrícico ("micotrófico"). Los gametofitos poseen morfología variada, útil para determinar la familia o al menos la clase a la que pertenecen: la mayoría de los helechos leptosporangiados poseen un gametofito con forma de corazón (como el de la foto), pero también hay gametofitos con forma elongada, filamentosa, o de otras formas. En el gametofito taloso se desarrollan las estructuras reproductoras multicelulares: arquegonios que darán las gametas femeninas inmóviles, y anteridios que darán anterozoides (gametas masculinas móviles flageladas). Los anteridios aparecen en la parte posterior del protalo, entre los rizoides, en los grupos primitivos están inmersos en el protalo y su pared no está diferenciada del resto del tejido excepto en la superficie, formando en su interior un elevado número de espermatozoides; en los grupos más recientes los anteridios toman la forma de pequeñas esferas compuestas por una pared dentro de la cual se forman las células espermatógenas que darán 16 o 32 espermatozoides; los anteridios poseen una célula llamada célula opercular, que es la que cede liberando los espermatozoides al exterior. Los arquegonios aparecen en la cara superior del gametofito, cerca de la escotadura, se forman a partir de una célula superficial que al dividirse va formando el cuello (única parte visible), el vaso y dentro de él la ovocélula y varias células más que degeneran formando una masa mucilaginosa. El mucílago atrae a los espermatozoides por quimiotaxis.
Al ser fecundado el oocito se formará el cigoto que por sucesivas divisiones mitóticas nutrido por el gametofito femenino, dará el embrión primitivo. El embrión se sigue desarrollando hasta adulto sin entrar en latencia. El embrión que proviene de un eusporangio es bipolar y con suspensor, posee meristema apical y meristema radical pero su raíz embrionaria nunca se desarrolla; el que proviene de un leptoesporangio es unipolar: solo posee el meristema apical.
La reproducción en pteridofitas heterosporadas
[editar]La descripción sobre el ciclo de vida dada hasta ahora se refiere a las familias de pteridofitos "isosporados", que tienen un "ciclo de vida isospórico". El nombre viene dado porque un mismo esporófito da esporas todas del mismo tamaño, que dan un solo tipo de gametofitos, que producen anteridios y arquegonios en el mismo talo. Algunas familias de pteridofitos siguen un ciclo de vida "heterosporado", esto es, producen dos morfos diferentes de esporas (esporas grandes o "megasporas" y esporas pequeñas o "microsporas"), que a su vez dan dos gametofitos diferentes (las megasporas producen gametofitos femeninos o arquegonios y las microsporas producen gametofitos masculinos o anteridios). A diferencia de lo que pasa con los helechos isosporados, y en forma similar a las espermatofitas, los gametófitos de los pteridofitos heterosporados se desarrollan por completo dentro de las paredes de la espora sin ser liberados nunca (por eso son "gametofitos endospóricos"). Lo que sí es liberado una vez madura, son los gametos masculinos (anterozoides), la gameta femenina se mantiene dentro de su arquegonio, el gametófito femenino se mantiene protegido por la pared de la megaspora mientras ocurre la fecundación y posterior desarrollo inicial del embrión.
Son heterosporados: los órdenes Selaginellales e Isoetales (que juntos forman un mismo clado —grupo monofilético con un ancestro común— en la división Lycophyta), y los helechos acuáticos Marsileaceae y Salviniaceae (que juntos forman el clado de "helechos heterospóricos" en la división Monilophyta). La heterosporía se desarrolló en forma independiente en estos dos clados.
Taxonomía y filogenia de pteridofitas
[editar]La hipótesis más aceptada hasta hace unos años sostenía que las pteridofitas eran un grupo monofilético (con un solo ancestro común) derivado de algún antecesor del alga verde. Hoy en día se sabe que esta división es un grupo parafilético que contiene un ancestro común a todas las plantas vasculares (que a su vez poseen un ancestro común con todas las briofitas). Se lo sigue estudiando por razones didácticas, como grupo contrapuesto a las espermatofitas (plantas con semilla donde la alternancia de generaciones se da en forma enmascarada).
Avances en filogenia
[editar]- Introducción teórica en Filogenia
En los últimos años se ha demostrado que Pteridophyta es un grupo parafilético que contiene dos líneas filogenéticas, Lycophyta y Monilophyta. Los últimos avances en filogenia (2013) dan el siguiente resultado:[6]
|
Pteridophyta |
Los nombres "Helechos y afines", "Plantas afines a los helechos", y "Helechos eusporangiados", así como el taxón "Pterophyta" y el mismo "Pteridophyta", son consideradas agrupaciones no monofiléticas de plantas según estudios anteriores, como en el siguiente cuadro:

Acerca de la filogenia de pteridofitas, todavía no hay consenso, pero el futuro se ve promisorio debido a los recientes avances en materia de análisis moleculares que, junto con los análisis clásicos de morfología, están permitiendo clarificar en mayor medida las relaciones entre los diferentes grupos.
Con respecto a eso, Pryer et al. (2001), basados en diferencias de ADN y morfológicas entre representantes de los diferentes grupos de pteridófitos, concluyen que:
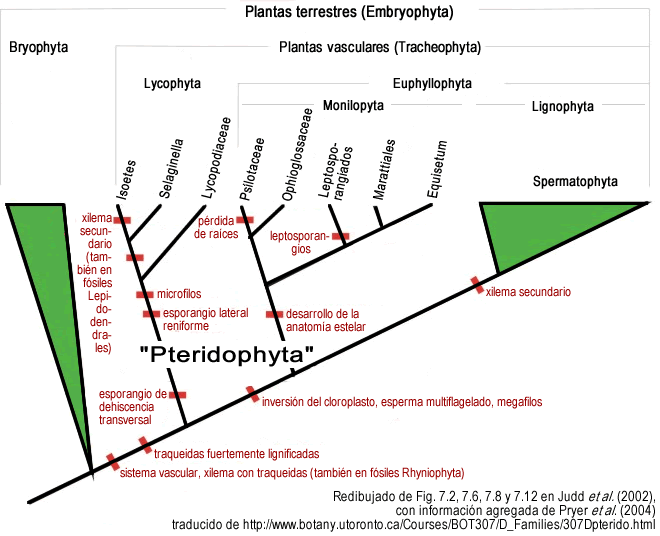
- Las divisiones Psilotophyta, Equisetophyta y Pterophyta constituyen un único grupo monofilético y son los parientes más cercanos a las espermatofitas (plantas con semilla), con lo que se descarta aparentemente la hipótesis de que los Psilotophyta y Equisetophyta sean grupos primitivos, o que sean eslabones evolutivos intermedios entre los briofitos y los Pterophyta (helechos con megafilos). Los primeros en sugerir la presencia de este grupo monofilético o "clado" fueron Kenrick y Crane (1997) basados en un carácter morfológico por el que llamaron al clado Infradivisión Moniliformopses (derivado del latín moniliformis: "forma de collar", debido a los lóbulos de protoxilema inmersos en el haz xilemático). Pryer et al. (2001) lo confirmaron como grupo monofilético aislado del resto de los clados, y posteriormente Judd et al. (2002) prefirieron darle el estatus de división y propusieron el nombre Monilophyta.
- En cambio la división Lycophyta se mantiene como grupo monofilético, pariente más lejano de las plantas con semilla y los demás grupos de pteridofitas.
- En conclusión, la publicación divide a las plantas vasculares en 3 grupos monofiléticos o "clados": 1) Lycopodiophyta, 2) Spermatophyta, y 3) Monilophyta (nombre usado hoy en día).
En otra publicación Archivado el 27 de agosto de 2010 en Wayback Machine. que hizo el grupo de investigación en el 2004 en el que hacen nuevos análisis moleculares de los taxones de monilophytas concluyen:
- Dentro de Monilophyta, reconocen a partir de varios estudios publicados por primera vez en 1995 y avalados por varios grupos de investigación, que la primera línea filogenética que difiere del resto es la que reúne a Psilotaceae con Ophioglossaceae, relación que se mantuvo en la oscuridad durante mucho tiempo debido a la extrema simplificación presente en el esporófito de las dos familias. Los caracteres morfológicos más importantes que permiten reunir a los dos grupos en una línea filogenética son: Ophioglossaceae tiene raíces simples, sin ramificación, sin pelos radicales, mientras que Psilotaceae directamente no tiene raíces; además los dos grupos tienen gametófitos axiales y subterráneos, esporangios ubicados en la cara adaxial, y desarrollo del esporangio del tipo eusporangiado, entre otros caracteres. Las evidencias moleculares parecían concluyentes a este respecto.
- Dentro de Monilophyta, los helechos leptosporangiados son reconocidos como un grupo monofilético, manteniéndose aparte de Equisetaceae y Marattiaceae (familia de helechos eusporangiados con frondes, cuya estela característicamente es una polistela).
Por lo que se avala la filogenia propuesta en el 2002 por Judd et al., con agregados. En resumen el siguiente árbol filogenético:
Visión histórica
[editar]Los distintos grupos de pteridofitas se han clasificado taxonómicamente de acuerdo con el siguiente resumenː
El término Pteridophyta fue concebido por Colm en 1872 a partir de Pteris (del griego "helecho", referido a un género de helecho) y equivale aproximadamente a Filices del Sistema de Jussieu (1774). De acuerdo con el sistema Eichler se clasificó del siguiente modo:[7]
- subreino A. Cryptogamae
- división III. Pteridophyta
- clase 1. Equisetinae (colas de caballo)
- clase 2. Lycopodinae (licopodios)
- clase 3. Filicinae (helechos)
- división III. Pteridophyta
Este sistema es similar al de Bessey (1907), que usó tres phylaː Calamophyta, Pteridophyta y Lepidophyta, equivalentes a las tres clases de pteridofitas de Eichler.
Engler, a su división Embryophyta asiphonogama, subdivisión Pteridophyta, añadió la cuarta clase Psilotopsida sensu Engler (1924), similar al moderno Sistema Kubitzki.
De los diferentes taxones en que pueden dividirse las pteridofitas sobresale el de los helechos, los cuales fueron llamados luego Pterophyta (Pteropsida si clase, a veces también como clase Filicatae), que son los pteridofitos con megafilos. Durante un tiempo se creyó que Pterophyta constituía una línea parafilética más reciente que los demás grupos de helechos sin megafilos, que eran llamados "grupos primitivos" y se creyó que algunos helechos sin megafilos (como los equisetos) los habrían perdido de forma secundaria (después de haberlos adquirido).
Los taxones con representantes vivos que se podían encontrar hasta hace poco (Smith 1955, Bold 1957, Zimmerman 1959 [1]) son los siguientes:
- División Psilotophyta (Psilotae si clase). Comprende a la familia actual sin hojas Psilotaceae. También se le llamó Rhyniophyta (Zimmerman 1976), pues se especulaba una relación con pteridofitas fósiles.
- División Lycopodiophyta, Lycophyta o Lepidophyta (Lycopodiatae si clase). Comprende a los taxones actuales Lycopodiaceae, Selaginella e Isoetes.
- División Equisetophyta, Sphenophyta o Calamophyta (Equisetatae si clase). Comprende al género actual Equisetum.
- División Pterophyta. (Pteropsida o Filicatae si clase). Comprende a todos los helechos con megafilos. Este taxón recibió una gran variedad de nombres a lo largo de la historia, incluyendo el mismo "Pteridophyta" y también "Filicopsida" (sensu Engler).
- Algunas clasificaciones modernizaron la ubicación de Ophioglossaceae (tradicionalmente ubicada junto a los "helechos verdaderos"), ubicándola en una división aparte (Ophioglossophyta) además de las 4 mencionadas (por ejemplo en wikispecies), clasificación que se acerca más a lo que se sabe hoy en día de sistemática de pteridofitas.
Hay que tener en cuenta que los estudios filogenéticos más recientes avalan a los sistemas de A. Smith 2006 y Christenhusz 2011.
Taxonomía
[editar]- Introducción teórica en Taxonomía
Christenhusz et al. 2011,[1][2][3] basada en Smith et al. 2006,[4] 2008;[5] que también provee una secuencia lineal de las licofitas y monilofitas.
Las divisiones y clases son las siguientes:
- Lycophyta (familias 1 a 3)
- Monilophyta ("helechos"), que contiene a las clases (Smith et al. 2006) o subclases (Christenhusz et al. 2011):
- Equisetopsida o Equisetidae (familia 4)
- Psilotopsida u Ophioglossidae (familias 5 y 6)
- Marattiopsida o Marattiidae (familia 7)
- Polypodiopsida o Polypodiidae (familias 8 a 48)
Importancia económica
[editar]La mayoría de las pteridofitas solo tienen importancia para los sistemáticos, siendo el uso económico más valorado el uso como ornamentales: los helechos arborescentes adornan los jardines botánicos tropicales y templados de todo el mundo, y muchas polipodiales se utilizan como plantas de hoja de interior, como los helechos serrucho y las especies de Adiantum, también son valoradas las diferentes especies llamadas "doradillas", y los helechos trepadores como Lygodium japonicum. El helecho acuático Ceratopteris es conocido por los acuaristas de todo el mundo. También hay helechos cuyas frondes se extraen para adornar ramos de flores (como Rumohra o Lophosoria quadripinnata[8]).
Las especies de Marsilea son fácilmente confundibles con tréboles de 4 hojas por los no iniciados, y a veces son vendidas como tales.
Salvinia es un helecho flotante que es conocido en Estados Unidos por taponar diques y crear un problema económico, actualmente se están estudiando sus enemigos naturales para intentar controlarlo mediante control biológico de plagas.
Azolla es un helecho flotante cultivado en los campos de arroz debido a su poder fertilizante. También a veces es considerado maleza.
En algunos lugares se comen los brotes tiernos de las hojas de diferentes especies de helechos. Por ejemplo, en ciertos lugares del sur de Chile se comen en ensalada los brotes de ampe o "perritos".[8][9]
Adiantum capillus-veneris posee actividad analgésica y antitusígena. Se emplea abundantemente en la España Mediterránea como remedio muy específico.[10]
Cultivo de helechos
[editar]El cultivo de estas plantas a través de tallos es el método de cultivo más común. Pero existen otros métodos de cultivo, como el cultivo por esporas, por ejemplo. A continuación se explicará el cultivo por esporas. Solo se necesita: esporas de helecho, tierra, lámina flexible de plástico transparente, agua y material orgánico de cualquier tipo (hojas, tallos, etc.).
Las esporas se obtienen raspando las hojas de los helechos que las contienen. Son los puntos negros o marrones que se encuentran debajo de algunas de sus hojas.
- En un tiesto o maceta, colocar el material orgánico entre la tierra del fondo y la tierra de arriba. Siendo la capa de tierra superior bastante fina.
- Esparcir las esporas por toda la superficie de la tierra de la maceta. Sin cubrirlas con más tierra.
- Esparcir agua en forma de gotas o "spray", sobre la tierra y las esporas.
- Finalmente, cubrir la maceta con la lámina plástica flexible, para evitar la evaporación del agua y retener el dióxido de carbono de que el material orgánico libera mientras se descompone. El dióxido de carbono es vital para el crecimiento de nuevos helechos, ya que los helechos aparecieron en una época donde en la atmósfera era común este gas.
Los helechos pueden tardar mucho en crecer, a veces más de 4 semanas, dependiendo de las condiciones donde se encuentre la maceta y la especie del helecho.
cuando empiecen a brotar, se debe perforar con una aguja el plástico que los cubre para que se vayan acostumbrando poco a poco a su nueva atmósfera, luego de brotar. Cuando crezcan lo suficiente, pueden ser trasplantados a cualquier jardín o maceta.
Véase también
[editar]- Desarrollo de los helechos
- Embriogénesis en pteridófitas
- Clasificación de los organismos vegetales
- Sistema de clasificación de monilophytas de Smith 2006
Taxones que comprenden a las pteridofitas:
Taxón superior que abarca a las pteridofitas y las espermatofitas:
Notas
[editar]- ↑ a b Christenhusz et al. 2011. A linear sequence of extant families and genera of lycophytes and ferns. Phytotaxa 19: 7-54.
- ↑ a b Preface to “Linear sequence, classification, synonymy, and bibliography of vascular plants: Lycophytes, ferns, gymnosperms and angiosperms” http://www.mapress.com/phytotaxa/content/2011/f/pt00019p006.pdf
- ↑ a b Corrections to Phytotaxa 19: Linear sequence of lycophytes and ferns http://www.mapress.com/phytotaxa/content/2011/f/pt00028p052.pdf
- ↑ a b c A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. "A classification for extant ferns". Taxon 55(3), 705-731 ( pdf)
- ↑ a b Smith, A.R., Pryer, K.M., Schuettpelz, E., Korall, P., Schneider, H., & Wolf, P.G. (2008) Fern classification, pp. 417–467 en: Ranker, T.A., & Haufler, C.H. (eds.), Biology and Evolution of Ferns and Lycophytes. Cambridge , Cambridge University Press.
- ↑ Theodor Cole & Hartmut Hilger 2013 Trachaeophyte Phylogeny Archivado el 4 de marzo de 2016 en Wayback Machine.
- ↑ Oudemans, C.A.J.A. (1896). Rangschikking der Planten. in Oudemans, C.A.J.A. & Hugo de Vries. Leerboek der plantenkunde, ten gebruike bij het hooger onderwijs. volume 2(2).
- ↑ a b Smith-Ramírez, Cecilia (1994). «Usos artesanales del bosque nativo. La extracción silenciosa». Revista Ambiente y Desarrollo X (2). p. 71-76 Artículo en línea. Archivado desde el original el 21 de octubre de 2007. Consultado el 17 de septiembre de 2021.
- ↑ Hoffmann Jacoby, Adriana (1999). El bosque chilote. Historia natural del archipiélago de Chiloé. Conservación y desarrollo sustentable de sus bosques y biodiversidad. Santiago: Defensores del Bosque Chileno. ISBN 956-7721-22-X.
- ↑ Rivera, D. et al. 1998. Manual de Teoría y Prácticas de Etnobotánica, Ed. Diego Marín. ISBN 84-95095-06-8
Bibliografía
[editar]General
[editar]- Cabrera, A.L. (Ed.) 1977. Flora de la provincia de Jujuy. Parte II: Pteridophyta. Colección Científica, Instituto Nacional de Tecnología Agropecuaria, Tomo XIII.
- Tryon, R.M. & A.F. Tryon. 1982. Ferns and allied plants with special reference to tropical America. Springer-Verlag, Nueva York.
- H. C. Bold, C.J. Alexopoulos & Th. Delevoryas (1989) Morfología de las Plantas y los Hongos. Omega, Barcelona, 911 pp., ISBN 84-282-0754-2
- Moran, R.C. 2003. Los Géneros de Helechos y Licofitas Neotropicales. Una guía para Estudiantes. Edición Especial para Sistemática de Plantas Tropicales (OET 2003-28). Organización para Estudios Tropicales.
- Elsa L. Cabral de Ferber, Daniel Cian. 2003. "Pteridófitos, guía de consultas". Editado por: Elsa L. Cabral de Ferber, Cátedra Diversidad Vegetal (Botánica II), Facultad de Ciencias Exactas y Agrimensura, Universidad Nacional del Nordeste, Argentina. (contenido en línea aquí)
- Ramón Palacios. 2006. "Pteridofita: guía y glosario". Cátedra Sistemática de Plantas Vasculares (Biodiversidad de tracheophyta), Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Buenos Aires, Argentina.
Sistemática
[editar]- Engler, A. 1926. Die natürlichen Pflanzanfamilien. 2º Ed.
- Engler, A. Prantl y continuadores. 1887-1915. Die Natürlichen Pflanzenfamilien. 23 tomos. Leipzig.
- Kenrick P, Crane PR. 1997. The Origin and Early Evolution of Land Plants. Smithsonian Institution Press, Washington, DC
- 2000. The PLANTS Database, database (version 5.1.1). National Plant Data Center, NRCS, USDA. Baton Rouge, LA 70874-4490 USA. http://plants.usda.gov
- Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (resumen en inglés aquí, pdf aquí).
- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition.Sinauer Axxoc, USA.
- Pryer, Kathleen M., Eric Schuettpelz, Paul G. Wolf, Harald Schneider, Alan R. Smith y Raymond Cranfill. 2004a. "Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences". American Journal of Botany 91:1582-1598 (resumen en inglés aquí Archivado el 26 de agosto de 2010 en Wayback Machine.).
- Pryer, K. M., Schneider, H, y Magallón, S. 2004b. "The radiation of vascular plants". En J. Cracraft y J. M. Donoghue (editores), Assembling the tree of life, 138-153. Oxford Univ. Press, New York.
Enlaces externos
[editar]Wikimedia Commons alberga una categoría multimedia sobre Pteridophyta.
Wikispecies tiene un artículo sobre Pteridophyta.
- Dr. David T. Webb, Courses, https://web.archive.org/web/20060911051251/http://www.botany.hawaii.edu/faculty/webb/
- Palacios-Rios, M. 2007. https://web.archive.org/web/20170107191324/http://www.helechos.com.mx/
Cursos introductorios a la diversidad de plantas y algas (en inglés).
- 2001-2006. Dra. Ma. Mercedes Arbo, Dra. Ana Maria González. "Hipertextos de Morfología de Plantas Vasculares". http://www.biologia.edu.ar/botanica/ En: J. S. Raisman, Ana M. González, Hipertextos del área de la Biología. http://www.biologia.edu.ar/
Sitio con textos muy completos y con muchas ilustraciones. En español, actualizado regularmente.
- Gran web de botánica de la Universidad de Extremadura lecciones hipertextuales de botánica Archivado el 8 de noviembre de 2010 en Wayback Machine.
- James D. Mauseth, Plant Anatomy Laboratory, Micrographs of plant cells and tissues, with explanatory text. http://www.sbs.utexas.edu/mauseth/weblab/ Sitio con fotos al microscopio de células y tejidos de plantas. (en inglés)
- Pryer's lab con los pdfs de las publicaciones en las que interviene la autora.
- P. F. Stevens, 2001 en adelante, Angiosperm Phylogeny Website. Versión 9, junio de 2008, y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/
- Gómez P., Luis D., 1971. Contribuciones a la Pteridología Costarricense en: Revista de Biología Tropical. (20)1: 31-43, 1972.
| Control de autoridades |
|
|---|
Datos: Q178249