พลาสติด
| พลาสติด | |
|---|---|
| การจำแนกชั้นทางวิทยาศาสตร์ | |
| โดเมน: | แบคทีเรีย Bacteria |
| ไฟลัม: | ไซยาโนแบคทีเรีย Cyanobacteria |
| เคลด: | Plastids Plastids |
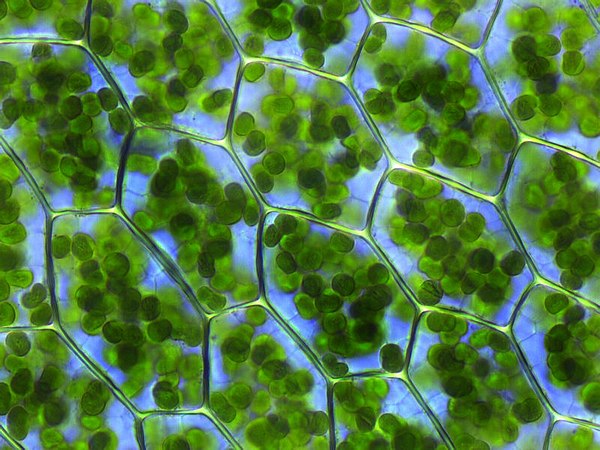
พลาสติด (อังกฤษ: plastid; กรีกโบราณ: πλαστός; plastos: ถูกก่อรูป) เป็นออร์แกเนลล์ที่มีเยื่อหุ้ม[1] พบใน เซลล์ของพืช, สาหร่าย, และยูแคริโอตอื่น ๆ พลาสติดเป็นไซยาโนแบคทีเรียร่วมอาศัยที่เกี่ยวข้องกับไซยาโนแบคทีเรียในสกุล Gloeomargarita[2] เหตุการณ์ที่นำไปสู่เอนโดซิมไบโอซิสแบบถาวรอาจเกิดขึ้นกับไซยาโนไบออนท์ (cyanobiont)[3] พลาสติดถูกค้นพบและตั้งชื่อโดย Ernst Haeckel แต่ผู้ที่ให้คำจัดความที่ชัดเจนเป็นคนแรกคือ A. F. W. Schimper พลาสติดเป็นแหล่งสร้างและสะสมสารประกอบที่สำคัญต่าง ๆ ในยูแคริโอตที่สร้างอาหารเองได้ มักบรรจุรงควัตถุที่ใช้ในการสังเคราะห์แสง ซึ่งประเภทของรงควัตถุในพลาสติดจะเป็นตัวกำหนดสีของเซลล์ พวกมันมีต้นกำเนิดร่วมกันและมีโมเลกุลดีเอ็นเอแบบเกลียวคู่ที่มีลักษณะเป็นวงกลมเช่นเดียวกับโครโมโซมแบบวงกลมของเซลล์โพรแคริโอต
พลาสติดในพืช
[แก้]
พลาสติดที่มีคลอโรฟิลล์สามารถสังเคราะห์ด้วยแสงได้และเรียกว่าคลอโรพลาสต์ พลาสติดสามารถเก็บสารผลิตภัณฑ์เช่น แป้ง และสามารถสังเคราะห์กรดไขมันและเทอร์พีน ซึ่งสามารถใช้ผลิตพลังงานและเป็นวัตถุดิบในการสังเคราะห์โมเลกุลอื่น ๆ ส่วนประกอบของคิวติเคิลและแว็กซ์ที่เคลือบผิวนอกของพืชถูกสังเคราะห์มาจากกรดปาลมิติกในอิพิเดอร์มัลเซลล์ ซี่งสังเคราะห์มาจากคลอโรพลาสต์ในเนื้อเยื่อมีโซฟิลล์อีกทีหนึ่ง[4]พลาสติดทั้งหมดพัฒนามาจากโพรพลาสติด ซึ่งมีอยู่ในบริเวณเนื้อเยื่อเจริญของพืช โพรพลาสติดและคลอโรพลาสต์อายุน้อยมักแบ่งตัวด้วยการแบ่งออกเป็นสอง กระทั่งคลอโรพลาสต์ที่เจริญมากกว่าก็มีความสามารถนี้เช่นกัน

ในพืช โพรพลาสติด (พลาสติดที่ยังไม่พัฒนาไปทำหน้าที่เฉพาะ) อาจพัฒนาไปได้หลายรูปแบบขึ้นอยู่กับหน้าที่ของพวกมันในเซลล์ โดยอาจพัฒนาเป็นรูปแบบดังต่อไปนี้:[5]
- คลอโรพลาสต์ : พลาสติดสีเขียวสำหรับการสังเคราะห์ด้วยแสง มีอิทิโอพลาสต์เป็นตัวตั้งต้น (precursor)
- โรโดพลาสต์: คลอโรพลาสต์สีแดงที่พบในสาหร่ายสีแดง
- มูโรพลาสต์ (ไซยาโนพลาสต์หรือไซยาเนลล์): คลอโรพลาสต์ของกลอโคไฟต์
- คลอโรพลาสต์ทุติยภูมิและตติยภูมิ: จากเอนโดซิมไบโอซิสของสาหร่ายสีเขียวและสาหร่ายสีแดง
- โครโมพลาสต์ : พลาสติดที่มีสี สำหรับการสังเคราะห์และจัดเก็บรงควัตถุ
- เจอรอนโทพลาสต์: ควบคุมการแยกชิ้นส่วนองค์ประกอบที่ใช้ในการสังเคราะห์ด้วยแสง ระหว่างกระบวนการเสื่อมสภาพตามอายุ
- ลิวโคพลาสต์: พลาสติดไม่มีสีที่สังเคราะโมโนทอร์พีน; ในบางครั้งลิวโคพลาสต์ปรับเปลี่ยนไปทำหน้าที่ที่จำเพาะยิ่งขึ้นดังต่อไปนี้:
- อะไมโลพลาสต์: สำหรับเก็บแป้งและตรวจจับแรงโน้มถ่วง (ในการเบนเนื่องจากแรงโน้มถ่วง)
- เอไลโอพลาสต์: สำหรับเก็บไขมัน
- โปรตีโนพลาสต์: สำหรับจัดเก็บและแปรรูปโปรตีน
- แทนโนโซม: สำหรับสังเคราะห์และผลิตแทนนินและโพลีฟีนอล
พลาสติดมีความสามารถที่จะเปลี่ยนรูปแบบไปทำหน้าที่อื่นหรือกระทั่งเปลี่ยนกลับไปมาได้หลายรูปแบบ นอกจากนี้เอพิโคพลาสต์ยังเป็นพลาสติดที่ไม่สังเคราะห์แสงของโพรโตซัวในไฟลัม Apicomplexa ที่ได้จากเอนโดซิมไบโอซิสทุติยภูมิ
พลาสติดแต่ละอันสามารถทำสำเนาของพลาสโตมที่มีขนาด 75–250 กิโลเบส จำนวนสำเนาของจีโนมต่อพลาสติดมีความแปรผัน โดยอาจเป็นได้ตั้งแต่ 1000 ในเซลล์ที่มีการแบ่งตัวอย่างต่อเนื่อง (ที่โดยทั่วไปมีจำนวนพลาสติดน้อยมาก) ไปจนถึง 100 หรือน้อยกว่าในเซลล์ที่เจริญเต็มที่แล้ว ซึ่งมีพลาสติดอยู่จำนวนมากเป็นผลจากการแบ่งแต่ละครั้ง พลาสโตมประกอบด้วยยีนประมาณ 100 ยีนที่เข้ารหัสไรโบโซมมัลอาร์เอ็นเอและทรานส์เฟอร์อาร์เอ็นเอ (rRNA และ tRNA) รวมทั้งโปรตีนที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสง, กระบวนการถอดรหัสและแปลรหัสยีนของพลาสติด อย่างไรก็ตาม โปรตีนเหล่านี้เป็นส่วนเล็กน้อยที่แสดงให้เห็นถึงโปรตีนที่จำเป็นต่อการสร้างและบำรุงรักษาโครงสร้างจนถึงการทำงานของพลาสติด ยีนในนิวเคลียสของเซลล์พืชเข้ารหัสของโปรตีนพลาสติดจำนวนมา และการแสดงออกของีนในพลาสติดและนิวเคลียสมีการกำกับร่วมกันเพื่อประสานงานให้การเจริญของพลาสติดที่เกี่ยวข้องกับการแบ่งเซลล์เป็นไปอย่างเหมาะสม
ดีเอ็นเอของพลาสติดอยู่ในรูปของสารเชิงซ้อนที่เกี่ยวพันกับเยื่อหุ้มชั้นใน เรียกว่า 'plastid nucleoids' แต่ละนิวคลีออยด์อาจมีดีเอ็นเอของพลาสติดมากกว่า 10 สำเนา โพรพลาสติดประกอบด้วยนิวคลีออยด์แห่งเดียวอยู่ตรงกลางของพลาสติด พลาสติดที่กำลังพัฒนามีนิวคลีออยด์จำนวนมากซึ่งอยู่ประจำที่ด้านข้างของพลาสติด ติดกับเยื่อหุ้มชั้นใน ระหว่างการพัฒนาโพรพลาสติดไปเป็นคลอโรพลาสต์และการพัฒนาจากพลาสติดชนิดหนึ่งไปเป็นชนิดอื่น นิวคลีออยด์มีการเปลี่ยนแปลงสัณฐาน, ขนาด, และตำแหน่งภายในออร์แกเนลล์นั้น ๆ การปรับรูปแบบของนิวคลีออยด์นี้เชื่อว่าเกิดขึ้นจากปรับเปลี่ยนองค์ประกอบและสัดส่วนของโปรตีนนิวคลีออยด์
พลาสติดหลายชนิดโดยเฉพาะอย่างยิ่งที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสงมีเยื่อหุ้มชั้นในหลายชั้น
ในเซลล์พืช ส่วนยาวบาง ๆ ที่ยื่นออกมาจากพลาสติดmujเรียกว่าสโตรมูล (stromule) อาจยื่นเข้าไปในไซโทพลาซึมและเชื่อมเข้ากับพลาสติดจำนวนหนึ่ง โดยโปรตีน (และอาจรวมถึงโมเลกุลขนาดเล็กอื่น ๆ) สามารถผ่านสโตรมูลนี้ได้ เซลล์ที่ได้จากการเพาะเลี้ยงส่วนมากมีขนาดใหญ่เมื่อเทียบกับเซลล์พืชทั่วไป เซลล์เหล่านี้มีสโตรมูลที่ยาว ทั้งยังมีจำนวนมาก ซึ่งขยายไปจนถึงรอบนอกของเซลล์
ใน ค.ศ. 2014 พบหลักฐานที่อาจเป็นการสูญเสียจีโนมของพลาสติดใน Rafflesia lagascae ซึ่งเป็นพืชดอกเบียน และใน Polytomella ซึ่งเป็นสาหร่ายสีเขียว โดยพืชทั้งคู่ไม่เกิดการสังเคราะห์ด้วยแสง และไม่พบยีนของพลาสติดแม้ว่าจะได้ค้นหาอย่างถี่ถ้วนแล้ว อย่างไรก็ตามข้อสรุปที่ว่าพลาสโตมของพืชทั้งสองหายไปทั้งหมดยังคงเป็นที่ถกเถียงกันอยู่[6] นักวิทยาศาสตร์บางคนยืนยันว่าการสูญเสียจีโนมของพลาสติดนั้นไม่น่าจะเกิดขึ้นได้เนื่องจากพลาสติดที่ไม่สังเคราะห์แสงยังมียีนที่จำเป็นต่อการสังเคราะห์ทางชีวภาพต่าง ๆ อยู่ เช่นการสังเคราะห์ทางชีวภาพของฮีม (heme)[6][7]
พลาสติดในสาหร่าย
[แก้]ในสาหร่าย คำว่าลิวโคพลาสต์ใช้สำหรับพลาสติดทั้งหมดที่ปราศจากรงควัตถุ หน้าที่ของพวกมันแตกต่างจากลิวโคพลาสต์ของพืช อิทิโอพลาสต์, อะไมโลพลาสต์, และโครโมพลาสต์มีเฉพาะในพืชและไม่มีในสาหร่าย[ต้องการอ้างอิง] พลาสติดในสาหร่ายและฮอร์นเวิร์ตยังอาจแตกต่างจากพลาสติดในพืชจากการที่มีไพรีนอยด์ (pyrenoid)
สาหร่ายกลอโคไฟต์มีมูโรพลาสต์ (muroplast) ซึ่งมีความคล้ายคลึงกับคลอโรพลาสต์ ทว่ามีผนังเซลล์เป็นเปบทิโดไกลแคน ที่คล้ายกับของโพรแคริโอต สาหร่ายสีแดงมีโรโดพลาสต์ (rhodoplast) ซึ่งเป็นคลอโรพลาสต์สีแดงที่ช่วยให้สามารถสังเคราะห์แสงได้ที่ระดับความลึกถึง 268 ม.[5] คลอโรพลาสต์ของพืชแตกต่างจากโรโดพลาสต์ของสาหร่ายสีแดงในด้านความสามารถในการสังเคราะห์แป้งซึ่งเก็บไว้ในรูปของแกรนูลภายในพลาสติด ในสาหร่ายสีแดง ฟลอริเดียนสตาร์ชจะถูกสังเคราะห์และเก็บไว้ในไซโทซอลนอกพลาสติด[8]
การถ่ายทอด
[แก้]พืชส่วนใหญ่สืบทอดพลาสติดจากฝั่งใดฝั่งหนี่ง โดยทั่วไปแล้วพืชดอก (angiosperm) จะสืบทอดพลาสติดจากเซลล์สืบพันธ์เพศเมีย ในขณะที่พืชเมล็ดเปลือย (gymnosperm) จำนวนมากได้รับพลาสติดจากเกสรเพศผู้ สาหร่ายยังสืบทอดพลาสติดจากฝั่งใดฝั่งหนี่งเช่นกัน ดังนั้นดีเอ็นเอของพลาสติดจากฝั่งที่ไม่ได้รับมาจึงหายไปอย่างสมบูรณ์
ในการผสมภายในชนิดเดียวกันตามปกติ (ส่งผลให้เกิดลูกผสมตามปกติของสปีชีส์หนึ่ง) การถ่ายทอดทางพันธุกรรมของดีเอ็นเอพลาสติดอาจได้มาจากฝ่ายใดฝ่ายหนึ่ง (uniparental) ทั้งสิ้น อย่างไรก็ตาม ในการสร้างลูกผสมระหว่างสปีชีส์จะพบว่าการถ่ายทอดไม่มีความแน่นอน แม้ว่าพลาสติดจะได้รับมาจากฝ่ายแม่เป็นส่วนใหญ่ในการสร้างลูกผสมระหว่างสปีชีส์ แต่ก็มีรายงานว่าพบลูกผสมของพืชดอกที่มีพลาสติดจากทางฝ่ายพ่อ ประมาณ 20% ของพืชดอก รวมทั้งแอลแฟลฟา (Medicago sativa) พบการถ่ายทอดพลาสติดที่ได้รับมาจากทั้งสองฝ่าย (biparental)[9]
ความเสียหายและการซ่อมแซมดีเอ็นเอ
[แก้]ดีเอ็นเอของพลาสติดจากต้นกล้าข้าวโพดอาจได้รับความเสียหายเพิ่มขึ้นเมื่อต้นกล้าพัฒนาต่อไป[10] ดีเอ็นเอได้รับความเสียหายในสภาพแวดล้อมออกซิเดชั่น (oxidative environment) ที่เกิดจากปฏิกิริยาโฟโตออกซิเดชั่น การสังเคราะห์ด้วยแสง และการถ่ายทอดอิเล็กตรอน โมเลกุลของดีเอ็นบางส่วนได้รับการซ่อมแซม ในขณะที่ดีเอ็นเอที่ไม่ได้รับการซ่อมแซมจะถูกย่อยสลายไปเป็นชิ้นส่วนที่ทำงานไม่ได้
โปรตีนซ่อมแซมดีเอ็นเอถูกเข้ารหัสโดยจีโนมจากนิวเคลียสของเซลล์ แต่สามารถย้ายตำแหน่งไปยังพลาสติดได้เพื่อรักษาเสถียรภาพและบูรณภาพของจีโนมโดยการซ่อมแซมดีเอ็นเอของพลาสติด[11] ตัวอย่างเช่นในคลอโรพลาสต์ของมอส Physcomitrella patens โปรตีนที่ใช้ในการซ่อมแซมดีเอ็นเอที่เข้าคู่ผิด (Msh1) ทำปฏิกิริยากับโปรตีนที่ใช้ในกระบวนการ recombinational repair (RecA และ RecG) เพื่อรักษาเสถียรภาพของจีโนมพลาสติด[12]
ต้นกำเนิด
[แก้]พลาสติดอาจเป็นไซยาโนแบคทีเรียเอนโดซิมไบโอติก เหตุการณ์เอนโดซิมไบโอติกหลักนี้ถูกตั้งสมมติฐานว่าเกิดขึ้นเมื่อประมาณ 1.5 พันล้านปีก่อน[13] และทำให้ยูแคริโอตสามารถสังเคราะห์ด้วยแสงโดยมีออกซิเจนเป็นตัวรับอิเล็กตรอนตัวสุดท้ายได้[14] สามเชื้อสายวิวัฒนาการได้ปรากฏขึ้นโดยมีชื่อเรียกที่แตกต่างกัน: คลอโรพลาสต์ในสาหร่ายสีเขียวและพืช, โรโดพลาสต์ในสาหร่ายสีแดง, และมูโรพลาสต์ในกลอโคไฟต์ พลาสติดเหล่านี้มีความแตกต่างกันทั้งในด้านรงควัตถุและโครงสร้างระดับจุลภาค ตัวอย่างเช่นคลอโรพลาสต์ในพืชและสาหร่ายสีเขียวสูญเสียไฟโคบิลิโซม (phycobilisome) ที่เป็นองค์ประกอบเก็บเกี่ยวแสงที่พบในไซยาโนแบคทีเรีย สาหร่ายสีแดงและกลอโคไฟต์ แต่กลับมีสโตรมาและกรานาไทลาคอยด์แทน กลอโคสโตไฟเชียนพลาสติดยังคงมีผนังเซลล์ที่หลงเหลือมาจากไซยาโนแบคทีเรีย ต่างจากคลอโพลาสต์และโรโดพลาสต์ พลาสติดปฐมภูมิทั้งหมดมีเยื่อหุ้มสองชั้น
ในทางตรงกันข้ามจากพลาสติดปฐมภูมิที่ได้จากกระบวนการเอนโดซิมไบโอซิสปฐมภูมิของไซยาโนแบคทีเรีย พลาสติดที่ซับซ้อนเกิดจากกระบวนการเอนโดซิมไบโอซิสทุติยภูมิ ซึ่งสิ่งมีชีวิตยูแคริโอตหนึ่งได้กลืนสิ่งมีชีวิตยูแคริโอตอื่นที่มีพลาสติดปฐมภูมิเข้าไป[15] หากยูแคริโอตหนึ่งกลืนกินสาหร่ายสีเขียวหรือสีแดงเข้าไปและยังคงรักษามันเอาไว้ได้ จะได้พลาสติดที่มีเยื่อหุ้มมากกว่าสองชั้น ในบางกรณีพลาสติดอาจถูกลดทอนความสามารถทางเมแทบอลึซึมและ/หรือการสังเคราะห์ด้วยแสง สาหร่ายที่มีพลาสติดซับซ้อนที่ได้จากเอนโดซิมไบโอซิสทุติยภูมิของสาหร่ายสีแดง ได้แก่สตรามิโนไพล์, แฮปโตไฟต์, คริปโทโมแนด, และไดโนแฟลเจลเลตเกือบทั้งหมด (=โรโดไฟต์) พวกที่มีเอนโดซิมไบโอซิสสาหร่ายสีเขียวได้แก่ยูกลีนิดและคลอราคนิโอไฟต์ (= คลอโรพลาสต์) เอพิคอมเพลกซาที่เป็นไฟลัมของโพรโทซัวตัวเบียนปรับไม่ได้ ซึ่งมีทั้งเชื้อก่อโรคมาลาเรีย (Plasmodium spp.), ทอกโซพลาสโมซิส (Toxoplasma gondii), และโรคอื่นทั้งในสัตว์หรือมนุษย์ ล้วนมีพลาสติดที่ซับซ้อน (แม้ว่าออร์แกเนลล์นี้จะหายไปใน apicomplexans บางชนิดก็ตาม เช่น Cryptosporidium parvum ซึ่งทำให้เกิด cryptosporidiosis) เอพิโคพลาสต์ที่แม้ว่าจะขาดความสามารถในการสังเคราะห์ด้วยแสงแล้ว แต่เป็นออร์แกเนลล์ที่จำเป็นและเป็นเป้าหมายที่สำหรับการพัฒนายาต้านเชื้อรา
ไดโนแฟลกเจลเลตและทากทะเลบางชนิด โดยเฉพาะอย่างยิ่งทากในสกุล Elysia ที่กินสาหร่ายเป็นอาหารและเก็บพลาสติดของสาหร่ายที่ย่อยแล้วเพื่อให้ได้ประโยชน์จากการสังเคราะห์ด้วยแสงของพลาสติดนั้น แต่หลังจากนั้นไม่นาน พลาสติดจะถูกย่อยในที่สุด กระบวนการนี้เรียกว่าเคลปโทพลาสที (kleptoplasty) จากภาษากรีก kleptes ; ขโมย
วงจรการพัฒนาของพลาสติด
[แก้]
ในปีค.ศ. 1977 JM Whatley ได้เสนอวงจรการพัฒนาของพลาสติด ซึ่งกล่าวว่าการพัฒนาพลาสติดไม่ได้เป็นแบบทิศทางเดียวเสมอไป หากแต่เป็นกระบวนการแบบวัฏจักรหลาย ๆ ครั้งมาประกอบกัน โพรพลาสติดเป็นตัวตั้งต้นของพลาสติดในรูปแบบที่แตกต่างกันมากขึ้นดังที่แสดงในแผนภาพดังแสดง[16]
Paulinella chromatophora
[แก้]พอลิเนลลามีออร์แกเนลล์ที่ใกล้เคียงพลาสติดที่ไม่ถูกจัดอยู่ในประเภทใดที่กล่าวมา เรียกว่าโครมาโตฟอร์ (chromatophore) ซึ่งเป็นออร์แกเนลล์ที่ได้รับมาจากการเอนโดซิมไบโอซิสเบตา-ไซยาโนแบคทีเรียเข้าไป[17] การเกิดเอนโดซิมไบโอซิสนี้เกิดขึ้นมาได้ไม่นานและยังเป็นรูปแบบที่สองที่ทราบของเอนโดซิมไบโอซิสปฐมภูมิของไซยาโนแบคทีเรีย
อ่านเพิ่ม
[แก้]อ้างอิง
[แก้]- ↑ Sato N (2006). "Origin and Evolution of Plastids: Genomic View on the Unification and Diversity of Plastids". ใน Wise RR, Hoober JK (บ.ก.). The Structure and Function of Plastids. Advances in Photosynthesis and Respiration. Vol. 23. Springer Netherlands. pp. 75–102. doi:10.1007/978-1-4020-4061-0_4. ISBN 978-1-4020-4060-3.
- ↑ Moore KR, Magnabosco C, Momper L, Gold DA, Bosak T, Fournier GP (2019). "An Expanded Ribosomal Phylogeny of Cyanobacteria Supports a Deep Placement of Plastids". Frontiers in Microbiology (ภาษาอังกฤษ). 10: 1612. doi:10.3389/fmicb.2019.01612. PMC 6640209. PMID 31354692.
- ↑ Vries, Jan de; Gould, Sven B. (2018-01-15). "The monoplastidic bottleneck in algae and plant evolution". Journal of Cell Science (ภาษาอังกฤษ). 131 (2): jcs203414. doi:10.1242/jcs.203414. ISSN 0021-9533. PMID 28893840.
- ↑ Kolattukudy, P.E. (1996) "Biosynthetic pathways of cutin and waxes, and their sensitivity to environmental stresses", pp. 83–108 in: Plant Cuticles. G. Kerstiens (ed.), BIOS Scientific publishers Ltd., Oxford
- ↑ 5.0 5.1 Wise, Robert R. (2006). "1. The Diversity of Plastid Form and Function". Advances in Photosynthesis and Respiration. Vol. 23. Springer. pp. 3–26. doi:10.1007/978-1-4020-4061-0_1. ISBN 978-1-4020-4060-3.
- ↑ 6.0 6.1 "Plants Without Plastid Genomes". The Scientist. สืบค้นเมื่อ 2015-09-26.
- ↑ Barbrook AC, Howe CJ, Purton S (February 2006). "Why are plastid genomes retained in non-photosynthetic organisms?". Trends in Plant Science. 11 (2): 101–8. doi:10.1016/j.tplants.2005.12.004. PMID 16406301.
- ↑ Viola R, Nyvall P, Pedersén M (July 2001). "The unique features of starch metabolism in red algae". Proceedings. Biological Sciences. 268 (1474): 1417–22. doi:10.1098/rspb.2001.1644. PMC 1088757. PMID 11429143.
- ↑ Zhang Q (March 2010). "Why does biparental plastid inheritance revive in angiosperms?". Journal of Plant Research. 123 (2): 201–6. doi:10.1007/s10265-009-0291-z. PMID 20052516. S2CID 5108244.
- ↑ Kumar RA, Oldenburg DJ, Bendich AJ (December 2014). "Changes in DNA damage, molecular integrity, and copy number for plastid DNA and mitochondrial DNA during maize development". Journal of Experimental Botany. 65 (22): 6425–39. doi:10.1093/jxb/eru359. PMC 4246179. PMID 25261192.
- ↑ Oldenburg DJ, Bendich AJ (2015). "DNA maintenance in plastids and mitochondria of plants". Frontiers in Plant Science. 6: 883. doi:10.3389/fpls.2015.00883. PMC 4624840. PMID 26579143.
- ↑ Odahara M, Kishita Y, Sekine Y (August 2017). "MSH1 maintains organelle genome stability and genetically interacts with RECA and RECG in the moss Physcomitrella patens". The Plant Journal. 91 (3): 455–465. doi:10.1111/tpj.13573. PMID 28407383.
- ↑ Ochoa de Alda JA, Esteban R, Diago ML, Houmard J (September 2014). "The plastid ancestor originated among one of the major cyanobacterial lineages". Nature Communications. 5: 4937. Bibcode:2014NatCo...5.4937O. doi:10.1038/ncomms5937. PMID 25222494.
- ↑ Hedges SB, Blair JE, Venturi ML, Shoe JL (January 2004). "A molecular timescale of eukaryote evolution and the rise of complex multicellular life". BMC Evolutionary Biology. 4: 2. doi:10.1186/1471-2148-4-2. PMC 341452. PMID 15005799.
- ↑ Chan CX, Bhattachary D (2010). "The Origin of Plastids". Nature Education. 3 (9): 84.
- ↑ Whatley, Jean M. (1978). "A Suggested Cycle of Plastid Developmental Interrelationships". The New Phytologist. 80 (3): 489–502. doi:10.1111/j.1469-8137.1978.tb01581.x. ISSN 0028-646X. JSTOR 2431207.
- ↑ Marin, Birger; Nowack, Eva CM; Glöckner, Gernot; Melkonian, Michael (2007). "The ancestor of the Paulinella chromatophore obtained a carboxysomal operon by horizontal gene transfer from a Nitrococcus-like γ-proteobacterium". BMC Evolutionary Biology. 7 (1): 85. doi:10.1186/1471-2148-7-85. PMC 1904183. PMID 17550603.
รายการอ่านประกอบ
[แก้]- Hanson MR, Köhler RH. "A Novel View of Chloroplast Structure". Plant Physiology Online. คลังข้อมูลเก่าเก็บจากแหล่งเดิมเมื่อ 2005-06-14.
- Wycliffe P, Sitbon F, Wernersson J, Ezcurra I, Ellerström M, Rask L (October 2005). "Continuous expression in tobacco leaves of a Brassica napus PEND homologue blocks differentiation of plastids and development of palisade cells". The Plant Journal. 44 (1): 1–15. doi:10.1111/j.1365-313X.2005.02482.x. PMID 16167891.
- Birky CW (2001). "The inheritance of genes in mitochondria and chloroplasts: laws, mechanisms, and models" (PDF). Annual Review of Genetics. 35: 125–48. doi:10.1146/annurev.genet.35.102401.090231. PMID 11700280. คลังข้อมูลเก่าเก็บจากแหล่งเดิม (PDF)เมื่อ 2010-06-22. สืบค้นเมื่อ 2009-03-01.
- Chan CX, Bhattacharya D (2010). "The origins of plastids". Nature Education. 3 (9): 84.
- Bhattacharya D, บ.ก. (1997). Origins of Algae and their Plastids. New York: Springer-Verlag/Wein. ISBN 978-3-211-83036-9.
- Gould SB, Waller RF, McFadden GI (2008). "Plastid evolution". Annual Review of Plant Biology. 59: 491–517. doi:10.1146/annurev.arplant.59.032607.092915. PMID 18315522. S2CID 30458113.
- Keeling PJ (March 2010). "The endosymbiotic origin, diversification and fate of plastids". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 365 (1541): 729–48. doi:10.1098/rstb.2009.0103. PMC 2817223. PMID 20124341.
แหล่งข้อมูลอื่น
[แก้]- Transplastomic plants for biocontainment (biological confinement of transgenes) — Co-extra research project on coexistence and traceability of GM and non-GM supply chains
- Tree of Life Eukaryotes เก็บถาวร 2012-01-29 ที่ เวย์แบ็กแมชชีน
- พลาสติด จากคณะเกษตรศาสตร์ มหาวิทยาลัยเชียงใหม่ เก็บถาวร 2010-05-27 ที่ เวย์แบ็กแมชชีน